
Эдвард Стил, Налин Чандра Викрамасингхе
Мы из космоса? Раз-/загадка Кембрийского взрыва
Кембрийским взрывом, как известно, называют период, когда внезапно, словно из ниоткуда на Земле появилось огромное разнообразие форм жизни, произошло резкое увеличение биоразнообразия скелетной фауны. Данные о Кембрийском взрыве основаны на очень ограниченных данных и его история является крайне дискуссионной в научных кругах. Даже Дарвин назвал этот период тем фактом, который не укладывается в его теорию постепенных эволюционных изменений. «Взрыв» был около 542 млн. лет назад и очень сложно отыскать его причину.
В 2018 году солидная международная научная команда под руководством австралийского иммунолога Эдварда Стила и при участии известного британского астробиолога Налина Чандры Викрамасингхе, опубликовала обширную обзорную статью, в которой в свете последних научных данных учёные попытались проанализировать гипотезу и обосновать несколько оригинальных тезисов, например, таких, как внеземная природа Кембрийского взрыва.
На метеоритах, упавших на Землю, периодически находят микробные окаменелости явно внеземного происхождения. А в прошлом году исследование микробов из космической пыли, налипшей на внешнюю сторону иллюминатора МКС показало: были выявлены жизнеспособные бактерии из типичных наземных и морских родов. Основываясь на множестве исследований авторы этой статьи выдвигают смелую гипотезу о том, что причиной Кембрийского взрыва – послужило прибытие на нашу планету вирусных генов из космоса, повлиявших затем на земную эволюцию и, в конечном счёте, появление человека.
Это кажется странным, но ведь были времена, когда и существование метеоритов отрицалось, а непосредственных свидетелей их падения поднимали на смех, просто потому что «камни не могут падать с неба, ведь на небе нет камней», как утверждал француз Антуан Лавуазье, основатель современной химии. Так что, кто бы что ни говорил, авторы настаивают: «Мы все родом из космоса».
Оглавление / Причина Кембрийского взрыва: земная или космическая?
-
Причина Кембрийского взрыва: земная или космическая?
Причина Кембрийского взрыва: земная или космическая?
Прогресс в Биофизике и Молекулярной Биологии 136 (2018) 3--23
|
|
Доступно на ScienceDirect ПРОГРЕСС В БИОФИЗИКЕ И МОЛЕКУЛЯРНОЙ БИОЛОГИИ Страница журнала: www.elsevier.com/locate/pbiomolbio |
|


Причина Кембрийского Взрыва: Земная или Космическая?
Эдвард Дж.Стилa,j ***, Ширван Аль-Муфтиb, Кеннет А.Августинc, Роана Чандраджитd, Джон П.Когланe, С.Дж.Кулсонb, Судипто Гошf, Марк Гилманg, Реджинальд М.Горджинскиh, Бриг Клайсb, Годфрей Луисi, Китсири Маханамаj, Кит Р.Оливерk, Хулио Падронl, Цзянвэнь Цюйm, Джон А.Шустерn, У.Е.Смитo, Дуэйн П.Снайдерb, Джулиан А.Стилp, Брент Дж.Стюартa, Роберт Тэмплq, Дженсуке Токороo, Кристофер А.Таутr, Александер Унзикерs, Милтон Уэйнрайтb,j, Джейми Уоллисb, Дэрил Х.Уоллисb, Макс К.Уоллисb, Джон Уэтераллt, Д.Т.Викрамасингхеu, Дж.Т.Викрамасингхеb, Н.Чандра Викрамасингхеb,j,o, Юншэн Люv,w
- *** a CY O'Connor ERADE Village Foundation, Пиара Уотерс, Вашингтон, Австралия
- b Букингемский центр астробиологии, Букингемский университет, Великобритания
- с Центр физики живых организмов, факультет физики, Мичиганский технологический университет, Мичиган, США
- d Департамент геологии, Университет Перадении, Перадения, Шри-Ланка
- e Мельбурнский университет, офис декана факультета медицины, стоматологии и медицинских наук, 3-й уровень, здание Алана Гилберта, Австралия
- f Институт металлургии и материаловедения, Канпур, Индия
- g Южноафриканский институт исследования мозга, 6 Campbell Street, Waverley, Йоханнесбург, Южная Африка
- h Сеть здравоохранения Университета Торонто, Общая больница Торонто, Университет Торонто, Канада
- i Кафедра физики, Кочинский университет науки и технологий, Кочин, Индия
- j Центр астробиологии, Университет Рухуна, Матара, Шри-Ланка
- k Школа ветеринарии и естественных наук Университета Мердока, Перт, Вашингтон, Австралия
- l Studio Eutropi, Клиническая патология и питание, Via Pompei 46, Ardea, 00040, Рим, Италия
- m Департамент по контролю за инфекционными заболеваниями, Тяньцзиньский центр по контролю и профилактике заболеваний, Китай
- n Школа истории и философии науки, факультет естественных наук, Сиднейский университет, Сидней, Австралия
- o Институт изучения панспермии и астробиологии, Гифу, Япония
- p Центр химии поверхности и катализа, KU Leuven, Celestijnenlaan 200F, 3001, Leuven, Бельгия
- q Фонд истории китайской культуры, Конвей-холл, Лондон, Великобритания
- r Институт астрономии, обсерватории, Мэдингли-роуд, Кембридж, CB3 0HA, Великобритания
- s Pestalozzi-Gymnasium, 11, D-81247, Мюнхен, Германия
- t Школа биомедицинских наук, Перт, Университет Кертина, Вашингтон, Австралия
- u Колледж физико-математических наук, Австралийский национальный университет, Канберра, Австралия
- v Хэнаньский совместный инновационный центр современного биологического разведения, Хэнаньский институт науки и технологий, Синьсян, 453003, Китай
- w Департамент биохимии, Университет Альберты, Эдмонтон, AB T6G 2H7, Канада
СПРАВКА О СТАТЬЕ
История статьи: Доступна онлайн с 13 марта 2018 года
Понятия: Космическая биология Кембрийский взрыв Ретровирусы
Панспермия Гипермутации и эволюция
Истоки эпидемий и пандемий
КРАТКИЙ ОБЗОР
В этой статье мы рассматриваем основные свидетельства, соответствующие гипотезе о Кометной (Космической) Биологии Хойла-Викрамасингхе (Х.-В.), а также те, которые были предугаданы ею. Многие из этих физических и биологических свидетельств многофакторны. Особое внимание уделяется недавним исследованиям, которые датируются моментом появления сложных ретровирусов у позвоночных во время, или непосредственно до Кембрийского взрыва, случившегося около 500 млн. лет назад. Известно, что такие вирусы, вероятно, могут быть связаны с основными эволюционными процессами генома. Мы считаем, что это совпадение не случайно, что согласуется с ключевым предположением теории Х.-В., согласно которому основные эволюционные границы вымирания-диверсификации на протяжении истории совпадали с явлениями учащенных столкновений с несущими вирусы кометными болидами. Второе направление
- удивительная эволюция усложнения интеллектуальной комплексности (головоногие моллюски), кульминацией которой стало появление осьминогов. Третий фокус внимания направлен на свидетельства присутствия в метеоритах окаменелых микроорганизмов наряду с обнаруженными в верхних слоях атмосферы видимыми частицами, несущими жизнь из космоса. На наш взгляд, совокупность многофакторных данных, собранных Фредом Хойлом, Чандрой Викрамасингхе и их многочисленными коллегами с 1960-х годов, приводит нас к весьма убедительному выводу: несущие жизнь кометы могли засеять семена жизни здесь, на Земле, в тот момент, как только условия на нашей планете позволили этим семенам начать прорастать (приблизительно 4,1 млрд. лет назад или чуть раньше); а живые организмы - такие как выносливые и резистентные к условиям космоса бактерии, вирусы, более сложные эукариотические клетки, оплодотворенные яйцеклетки и семена, которые с тех пор непрерывно доставлялись на Землю, - таким образом явились одним из основных факторов дальнейшей земной эволюции, которая привела к значительному генетическому разнообразию и появлению человечества.
1 ©2018 Авторы. Опубликовано Elsevier Ltd. Это статья открытого доступа. Лицензия CC BY-NC-ND (http://creativecommons.org/licenses/by-nc-nd/4.0/).
СОДЕРЖАНИЕ
- Цель статьи
- Вводные замечания
- Космическая теория жизни
- Условия наличия жидкой воды
- Самая ранняя земная жизнь
- Происхождение жизни
- Органические молекулы и биологическая пыль в космосе и в кометах
- Доказательство существования жизни на Марсе
- Микробный материал в стратосфере и в метеоритах
- Принципы вирусологии - вирусы как плотные информационно-насыщенные системы контроля
- Критика особенностей хозяина-носителя вирусов и его выживаемость в космических условиях
- Ретровирусный индукционный режим
- Эволюция усложнения интеллектуальной комплексности
- Эволюционное происхождение ретровирусов позвоночных
- Кембрийский взрыв
- Последствия Кембрийского взрыва и данные о ретровирусах
- Космическая биология и зарождение человечества
- Причины эпидемий и пандемий: новый принцип вирусологии
- Смена парадигмы
- Конфликты интересов
- Вклад авторов
- Примечание к доказательству
- Происхождение жизни и научные альтернативы космической панспермии Х.-В.?
- Ретровирусы, ретроэлементы Alu и LINE: структура, функции, подвижность генома и значение
- Анализ причин и движущих сил истории происхождения жизни на Земле, изложенных в этой статье
- Ссылки
“У историка, занимающегося изучением науки, может возникнуть соблазн заявить, что когда меняются парадигмы, вместе с ними меняется и сам мир”. Структура научных революций (Thomas S Kuhn, 1962 , 2nd ed.1970)
Когда ему предоставили описания необъяснимых характеристик выживания Тихоходок, друг воскликнул: “Как они вообще эволюционировали?” (Аноним, 2017)
“Идея о том, что во всей Вселенной жизнь существует лишь для Земле, по существу восходит ко временам до Коперника. Опыт уже неоднократно научил нас, что такой тип мышления, вероятнее всего, ошибочен. Почему лишь наша собственная, бесконечно малая ниша во Вселенной должна быть уникальной? Точно так же, как ни одна страна не является центром Земли, так и Земля не является центром Вселенной”. Облако жизни (Fred Hoyle and N Chandra Wickramasinghe, 1978, J.M.Dent & Sons, London, p.132).
- Цель статьи
Цель этой аналитической статьи заключается, в основном, в обзоре коллективных знаний и опыта более 30 ученых и исследователей ряда дисциплин физических и биологических наук. Мы рассмотрели большую часть собранных за последние 60 лет ключевых экспериментальных и эмпирических данных, соответствующих идеям гипотезы Хойла-Викрамасингхе по Кометной (Космической) Биологии, или спрогнозированных ею.
Мы четко осознаем, что распространенное мнение о происхождении и дальнейшей эволюции жизни на Земле прочно укрепилось за “Земной” парадигмой. Наша цель - побудить представителей научных кругов из областей биофизики и биомедицины, а также сообщества ученых, занимающихся вопросами эволюции, к дальнейшему обсуждению совершенно иной точки зрения о “космическом” происхождении жизни, предложенной Хойлом и Викрамасингхе. На наш взгляд, их теория включает в свой анализ гораздо более обширный спектр физических, астрофизических, биологических и биофизических явлений, которые часто не только совершенно невозможно объяснить, но которые противоречат господствующей земной неодарвиновской парадигме. Более того, если некоторые читатели надеются найти здесь исследование, основанное на анализе популяционной генетики, как это прокомментировал один из рецензентов: “...анализ темпов эволюции, примеры появления новых генов, не имеющих гомологии со старыми, и т.д.”, то они во многом будут разочарованы, хотя согласно полученным недавно данным, некоторые генетические характеристики осьминогов и других головоногих моллюсков представляют собой сложные примеры традиционного эволюционного мышления. Но это не основное направление нашего анализа.
Общепринятая, и безусловно необычная, особенность научной работы характеризуется тем, что необходимо обеспечить понятное общение на доступном английском языке между разнообразными научными дисциплинами. Однако, учитывая специфический интерес, мы обсуждаем недавние филогенетические данные о сложных ретровирусах у линии позвоночных во время или незадолго до Кембрийского взрыва, произошедшего около 500 млн. лет назад (широко признанное эпохальное событие в эволюционной истории многоклеточной жизни на Земле). Предполагается, что эти типы вирусов с обратной транскрипцией2 и интеграцией генома могут быть связаны с основными эволюционными геномными процессами. Мы считаем, что совпадение с Кембрийским взрывом является, вероятно, не случайным, и, вместе с тем, оно согласуется с ключевым предположением гипотезы Х.-В., согласно которому основные эволюционные границы вымирания-диверсификации совпадают с явлениями учащенных столкновений с кометными болидами, несущими и доставляющими на Землю гипотетические вирусы, микроорганизмы и более сложные эукариотические системы на протяжении последних 4,5 млрд. лет истории Земли. Необязательно, что все доставленные таким образом живые системы смогли удержаться на нашей планете. Подразумевается, что существенные эволюционные процессы на самой Земле (какими бы ни были фактические молекулярно-генетические механизмы) также продолжали происходить своим чередом.
По нашему твердому убеждению, совокупность многофакторных данных и критического анализа, собранных Фредом Хойлом, Чандрой Викрамасингхе и их многочисленными коллегами, приводит к самому начальному, но убедительному научному выводу о том, что несущие жизнь кометы могли засеять семена жизни здесь, на Земле, в тот момент, как только условия на нашей планете позволили этим семенам начать прорастать (приблизительно 4,1 млрд. лет назад или чуть раньше); а живые организмы - такие как выносливые и резистентные к условиям космоса бактерии, вирусы, более сложные эукариотические клетки и организмы (например, тихоходки) и даже оплодотворенные яйцеклетки и семена растений, - возможно, с тех пор непрерывно доставлялись на Землю, помогая стимулировать дальнейший прогресс земной биологической эволюции. Со времен Лорда Кельвина (1871) и Сванте Аррениуса (1908) этот процесс носит научное название “Панспермия”.
Возможно, самые важные астрономические данные, имеющие в последние годы отношение к появлению теории о космической жизни, связаны с обнаружением обитаемых экзопланет - планет за пределами нашей солнечной системы. Общее количество таких, похожих на Землю планет только в нашей галактике Млечный Путь сейчас составляет около 100 миллиардов, а с учетом примерно 100 миллиардов галактик в наблюдаемой Вселенной, общее количество составляет 1022. Поскольку обмен возможно плодородным материалом между соседствующими местами обитания более чем вероятен, можно утверждать, что панспермия и теория космической жизни стали неизбежной данностью.
2Процесс образование двуцепочечной ДНК на основании информации в одноцепочечной РНК, при котором РНК-зависимая ДНК-полимераза катализирует синтез ДНК на матрице РНК. (Прим.пер.)
Сдвиг парадигмы к этой критической точке зрения, хотя и происходит, но еще отнюдь не завершен, поэтому мы полагаем, что сейчас настал исторический момент для всестороннего и продуманного междисциплинарного обзора значительной части соответствующих доказательств, которые и будут изложены в этой статье. Это новое научное осознание повлечет за собой много далеко идущих последствий, изучение которых, как мы считаем, станет привилегией будущих поколений.
- Вводные замечания
Аристотелевская парадигма спонтанного зарождения жизни - идея о том, что простейшие формы жизни возникли на Земле спонтанно (светлячки - из смеси теплой земли и утренней росы), - в той или иной форме просуществовала более 2000 лет. За это время она неоднократно выдерживала противостояние с противоречащими ей данным. Эксперименты Пастера в 1862 году по ферментации вина и сквашиванию молока побудили его сформулировать изречение “Omne vivum ex vivo” или “Вся жизнь происходит из жизни”. Смысл эксперимента Пастера заключался в том, что каждому поколению каждого микроба, растения или животного предшествовало поколение одного и того же организма. Эта точка зрения была с энтузиазмом поддержана другими, особенно физиками, и в частности одним из них, выдающимся Джоном Тиндаллом, который 21 января 1870 года прочел лекцию в Королевском Институте в Лондоне о значении панспермии. Примечательно и интересно, что учрежденный накануне журнал Nature3 рьяно возражал против этой лекции в своих редакционных колонках. Противостояние было продиктовано пониманием того, что в случае, если изречение Пастера является чистой правдой, то истоки жизни следует искать за пределами Земли. Дальнейший антагонизм к сформулированному Пастером значению панспермии проложил путь к появлению доминирующей биологической парадигмы - абиогенеза в первичном бульоне4. Последняя идея была разработана в то время, когда самые ранние живые клетки считались чрезвычайно простыми структурами, которые впоследствии могли развиться Дарвиновским путем. Конечно, после открытия чрезвычайно сложных молекулярных структур строения белка и ДНК, эти идеи надлежало критически проверить и опровергнуть. Но этого не произошло. Современные идеи абиогенеза в гидротермальных источниках срединно-океанических хребтов или где-либо еще на первобытной Земле превратились в изощренные предположения, практически не имеющие доказательной базы.
3Журнал “Природа” (Прим.пер.)
4Абиогенез - теория возникновения живого из неживого.
Первичный бульон - термин, введенный советским биологом А.И.Опариным, и использованный в выдвинутой им в 1924 году теории о возникновении жизни на Земле через превращение - в ходе постепенной химической эволюции содержащих углерод молекул - в первичный бульон. (Прим.пер.)
Даже если мы согласимся, что доминирующая неодарвиновская парадигма естественного отбора может объяснить аспекты эволюционной истории жизни после того, как жизнь зародилась, независимый абиогенез в космологически крошечном масштабе океанов, озер и гидротермальных источников срединно-океанических хребтов остается гипотезой, не имеющей практического подтверждения, и, более того, является лишним и ненужным. С учетом астрономических данных, указывающих на существование сотен миллиардов пригодных для жизни планет только в нашей галактике (Abe et al., 2013; Kopparapu, 2013), такой вариант поиска независимых источников жизни на какой-либо только одной планете, кажется, больше не является необходимым. Недавнее исследование засвидетельствовало наличие микробной жизни в канадских горных породах, образовавшееся 4,1-4,23 миллиарда лет назад (Dodd et al., 2017), и, на наш взгляд, если принять его во внимание, это затруднит рассмотрение возможности абиогенеза, имеющего место где-либо на Земле. Утверждения о том, что эти породы могли быть связаны с гидротермальными источниками срединно-океанических хребтов, все еще поднимает вопрос о том, как могла возникнуть жизнь на местах в эпоху раннего Катархея5, для которой были характерны частые и жесткие столкновения с астероидами и кометами. Скорее, мы считаем более разумным предположить, что конкретное доказательство наличия микробной жизни в канадских горных породах было доставлено кометными болидами только для того, чтобы быть мгновенно уничтоженным или карбонизированным при ударе.
Температурные условия, которые, скорее всего, преобладали вблизи изрешеченной ударами земной поверхности 4,1-4,23 млрд. лет назад, были слишком горячими, чтобы выжить, даже для простых органических молекул, не говоря уже о том, чтобы эволюционировать в живую сложную структуру. Это выдвигает панспермию в качестве наиболее вероятного и обоснованного варианта происхождения земной жизни: скорее всего, первые микробы были доставлены на планету вместе с кометами и метеоритами. Этот анализ, наряду с результатами работы Белль и ее коллег (Bell and associates, 2015), датирующих цирконы из Джек-Хиллса в Западной Австралии аналогичным периодом времени, представляет собой самое последнее открытие и, как результат, естественным образом ведет к идеям, которые мы подробно обсуждаем в этой статье.
С приходом 20го века противостояние панспермии еще глубже укоренилось в нашей научной культуре. Попытки Хойла и Викрамасингхе (Hoyle and Wickramasinghe, 1979, 1981, 1986, 1993; Wickramasinghe, 2015a, 2015b) пересмотреть и восстановить панспермию в свете новых данных в областях астрономии и биологии часто встречались враждебно (Hoyle and Wickramasinghe, 1986; Wickramasinghe, 2015a).
Подобная судьба часто постигала попытки восстановить некоторые важные аспекты ламаркизма, додарвиновского представления о том, что гены в нашем геноме могут быть
5 Интервал геологического времени, предшествовавший Архею, начавшийся примерно 4,54 млрд. лет назад. (Прим. пер.)
“направленно” обогащены посредством наследования адаптивных, обусловленных экологией и приобретенных характеристик (Steele, 1979; Steele et al., 1998; Jablonka and Lamb, 1995, 2005; Lindley, 2010). Этот последний механизм генного наследования можно более точно описать как обратное проникновение между сомой и зародышевой линией через полупроницаемый (не абсолютный) Барьер Вайсманна, концепцию, появившуюся в 19м веке, примерно в период смерти Дарвина. В настоящее время имеется значительное количество доказательств, согласующихся с исходной “гипотезой соматического отбора” (предложенной одним из соавторов этой статьи (E.J.S.)6 в конце 1970-х годов), которая, как предполагается, действует через управление эндогенными ретровирусными векторами и обратную транскриптазу (Steele, 1979). Более того, Коррадо Спадафора, Йошенг Лю, Денис Ноубл, Джон Маттик и другие встали во главу согласующегося с этой позицией ряда современных дискурсов, наблюдений и критического анализа о том, что такие разработки в эволюционной биологии и смежных областях, как процессы наследования Ламарка (как прямые модификации ДНК, так и косвенные, а именно эпигенетические, трансмиссии) теперь требуют полного пересмотра стандартной неодарвиновской теории эволюции или “Нового синтеза”, появившейся в 1930-х и 1940-х годах (https://royalsociety.org/science-events-and-lectures/2016/11/evolutionary-biology/; Spadafora, 2008; Liu, 2007; Noble 2011; Noble 2013; Noble et al., 2016; Mattick, 2012; Liu and Li, 2016a; Liu and Li, 2016b). Помимо этого, много лет назад на основе имеющихся на тот момент сведений и наличия ряда внутренних противоречий в используемых данных (Hoyle and Wickramasinghe, 1979, 1981, 1982; Steele, 1979) этот призыв прозвучал от двух соавторов этой статьи (E.J.S., N.C.W.7).
Мы определенно не хотим, чтобы эту статью считали, как выразился один рецензент:“...чем-то вроде последней отчаянной попытки убедить основой поток научного сообщества, что, следуя неодарвинизму, они серьезно сбились с пути, потому что жизнь была перенесена на эту планету откуда-то из Вселенной на кометах/метеоритах и не является результатом абиогенеза на Земле”. На самом деле, мы считаем, что определенные механистические аспекты неодарвинистического и популяционно-генетического мышления имеют неоценимое значение в биомедицинских исследованиях и клинической медицине (разработка алгоритма “больших данных”, которые позволяют “персонализировать” навигацию по генетическим особенностям тысяч геномов человека, как, например, это делается в Broad Institute в Бостоне и в Wellcome Trust Sanger Institute в Кембридже). Однако, по нашему мнению, эти базовые дарвиновские концепции должны быть отграничены и помещены в космические, а не исключительно земные, условия. Таким образом, на наш взгляд, и панспермия, и вопросы Ламарка способствуют нашему
6 E.J.S. здесь и далее: Эдвард Дж.Стил - австралийский молекулярный иммунолог, изучающий вирусологию и эволюцию (Прим.пер.)
7 N.C.W. здесь и далее: Нали́ н Ча́ндра Викрамаси́ нгхе - британский физик, астроном и астробиолог; директор “Кардиффского центра Астробиологии”; совместно с Ф.Хойлом разработал гипотезу панспермии. (Прим.пер.)
более широкому пониманию, поскольку они затрагивают самую суть того, как возникла жизнь на Земле и как впоследствии она развивалась и диверсифицировалась до более высоких уровней сложного многообразия, которое мы наблюдаем сегодня. С нашей точки зрения, “естественный отбор” по своей сути (выживание наиболее приспособленных) все еще играет решающую роль в изменяющейся окружающей среде, но теперь он происходит в космических, а не в чисто земных условиях; и это согласованно происходит как с недарвиновскими, так и с неменделеевскими механизмами наследования. Тем не менее, мы признаем, что вся тема “эволюционных механизмов”, как и политических убеждений, одновременно является небезопасной и подогретой зоной социальных и культурных разногласий, и, безусловно, находится во всех тех областях, которые лежат за пределами нормального научного исследования. Тем не менее, простой факт, который нельзя отрицать, заключается в том, что земная биосфера является бесконечно малой частью гораздо, гораздо большей системы, чем космос астронома, и эти две системы неразрывно связаны.
Таким образом, мы со всей серьезностью предупреждаем: хотя точная научная терминология может иногда “коробить слух” по причине своей предыстории, но все же иногда ее применение является неизбежным, поэтому мы будем использовать такие понятия, как панспермия и ламаровское наследие, в нашей статье там, где сочтем это целесообразным, несмотря на их эмоциональное воздействие и предвзятую окраску.
Еще один наш аргумент в пользу возрождения этих безусловно противоречивых вопросов заключается в следующем: неправильные теории попросту не приносят плодов. Однако правильные теории всегда приводят логическими путями к последующим подтверждающим примерам и прогнозируют откртытие оригинальных явлений в реальном мире, другими словами, они не опровергают себя же суровыми экспериментальными и обсервационными8 испытаниями, выходящими за пределы их непосредственной разъяснительной области. Поэтому в настоящей статье мы именно в таком духе обсуждаем также захватывающие новые вирусологические данные, недавно опубликованные П.Эюсаканом и А.Кацуракисом (Aiewsakun and Katzourakis, 2017) в Nature Communications9 , которые подтверждают важный прогноз теории космической биологии Хойла-Викрамасингхе (Х.-В.) в деле защиты величайшего эпохального эволюционного события на Земле - Кембрийского взрыва многоклеточной жизни полмиллиарда лет назад.
Однако, прежде чем перейти к деталям, мы хотим подготовить почву для лучшего понимания открытия Эюсакана-Кацуракиса, обсудив важные экспериментальные и обсервационные данные, лежащие в основе концепции Х.-В. о повсеместном присутствии
8 Обсервационное исследование (наблюдательное исследование) - клиническое исследование, в котором исследователь собирает данные путем простого наблюдения событий в их естественном течении, не вмешиваясь в них активно.
9 Научный журнал “Общение Природы” (Прим.пер.)
космической биологии, непрерывно влияющей на Землю, и другие общие эволюционные проблемы, а также вирусные частицы и их общие свойства.
- Космическая теория жизни
В середине 1970-х годов идея существования пребиотических молекул в межзвездном пространстве или в кометах поначалу не являлась частью распространенного научного мнения. В 1974 году один из соавторов этой статьи (N.C.W.) вынес исходное предположение о присутствии органических полимеров в межзвездном пространстве (Wickramasinghe, 1974), за которым последовала длинная серия статей при сотрудничестве с Фредом Хойлом и, как результат, прямое противостояние с господствующей научной парадигмой о происхождении жизни на Земле, так называемой теорией Первичного бульона Холдейна-Опарина (Hoyle and Wickramasinghe, 1976, 1977а, 1977b, 1978а, 1978b). После обсуждения ряда всевозможных условий в межзвездном и околозвездном пространстве, необходимых для зарождения биохимии (пребиотическая эволюция), Хойл и Викрамасингхе (Hoyle and Wickramasinghe,1978b) остановились на кометном ансамбле, состоящем из примерно 1011 комет в одной только нашей солнечной системе, как предпочтительной среде для зарождения жизни на Земле (Hoyle and Wickramasinghe, 1985, 1986). Утверждалось, что предполагаемые радиоактивно подогретые внутренние части этих ледяных тел, которые содержат области с жидкой водой (см.ниже), изобилующие межзвездной органикой, являются намного более благоприятными для происхождения жизни, чем любые из условий на Земле.
- Условие наличия жидкой воды
Наличие жидкой воды является непременным условием не только для возникновения жизни, но и для активных микробиологических процессов. Это условие вышло на первый план и получило широкую огласку после недавнего завершения миссии НАСА “Кассини”. Когда Фред Хойл и один из соавторов этой статьи (N.C.W.) впервые выдвинули и развили теорию о кометной панспермии (Hoyle and Wickramasinghe, 1979, 1981, 1985), прямых доказательств о существовании жидкой воды где-либо, помимо Земли, еще не существовало. Вывод о наличии жидкой воды в кометах и гигантских ледяных телах Солнечной системы был сделан только на основании теоретических исследований. Хойл и Викрамасингхе доказывали (Hoyle and Wickramasinghe, 1985), что, как правило, ледяное тело, состоящее из естественной для солнечной системы фракции урана и тория, за счет радиоактивного нагрева будет поддерживать теплые внутренние океаны жидкой воды, обеспечивая таким образом среду обитания для микробов в течение миллиардов лет. Хойл и Викрамасингхе писали (Hoyle and Wickramasinghe, 1985): “Очевидно, что для тела
лунного размера R>1000 км поддержание жидкого состояния внутри своего тела не составит никакого труда, а некоторые кометы, возможно, смогли бы сделать это в течение, по крайней мере, первых 500 миллионов лет истории Солнечной системы. Избыточный выход энергии просто приведет к утончению оболочки поверхности, тогда как уменьшение мощности приведет к утолщению оболочки. По сути дела, толщина оболочки будет регулироваться в зависимости от мощности выпускаемой энергии. Это решает проблему существования хемоавтотрофных биологических систем в анаэробных условиях.” Позже Н.Ч. Викрамасингхе (Wickramasinghe et al.1996) и Дж.Т. Викрамасингхе (Wickramasinghe et al. 2009) провели более подробные исследования тех же процессов. Спустя долгое время в результате исследования космоса были окончательно установлены прямые доказательства наличия жидкой воды в кометах, а также в других ледяных телах Солнечной системы.
- Самая ранняя земная жизнь
Три десятилетия назад считалось, что самым ранним свидетельством микробной жизни в геологической летописи являются окаменелости, похожие на цианобактерии, которые появились 3,5 миллиарда лет (Ga)10 назад. С момента образования на Земле устойчивой коры около 4,3 млрд. лет назад, сразу после периода жестких столкновений с кометами (Эпоха Катархея, о которой мы уже упоминали), имелся временной промежуток в 800 млн.лет, в течение которого мог сформироваться канонический первичный бульон Холдейна-Опарина. Однако, недавние открытия показали, что этот временной интервал был практически закрыт (Dodd et al., 2017). Кроме того, было обнаружено, что обломочные цирконы ≥14 млрд.лет, которые были найдены в породах, принадлежащих к геологическому обнажению в районе Джек-Хиллз в Западной Австралии, содержат графитовые сферы микронных размеров со следами изотопов биогенного углерода (Bell et al., 2015). Следовательно, 12C-Обогащение, обнаруженное в этих вкраплениях, может рассматриваться как убедительное недвусмысленное доказательство существования микробной жизни на Земле более чем 4.1 млрд.лет назад, в эпоху столкновения комет и астероидов. Ввиду только что рассмотренных данных о ранних признаках клеточной жизни (≥4,1 млрд.) в осадках старейших гидротермальных источников в Канаде, эта информация также совпадает с датами появления первых признаков жизни на Земле (Dodd et al., 2017). Следуя ортодоксальному абиогенному мышлению, теперь требуется, чтобы произошло практически мгновенное преобразование неживого органического вещества в бактериологическую субстанцию, - мы полагаем, что такое предположение запредельно подрывает доверие к ограниченному лишь пределами Земли абиогенезу. Гораздо более правдоподобная вероятность состоит в том, что полностью развитые микроорганизмы и, возможно, другие эукариотические организмы прибыли на Землю на сталкивающихся с ней кометах, а позже они карбонизировались и оказались в ловушке конденсирующих минеральные зерна конгломератов. Сейчас становится убедительно ясно, что существуют сотни миллиардов планет земного типа и других благоприятных для жизни планетных тел,
10 Ga, от Giga-Annum, - гигагод, единица измерения времени, равная одному миллиарду лет. (Прим.пер.)
и обмен материалами между ними (метеоритами, кометными болидами) должен регулярно иметь место (Wickramasinghe et al., 2012; Kopparapu, 2013; Приложение А на стр. 40). Поэтому, с нашей точки зрения, кому-то придется сделать вывод о том, что вся галактика (и, возможно, наша локальная группа галактик) составляет единую связанную биосферу.
- Происхождение жизни
Поверхностная критика, которая часто выдвигается против теории космической жизни, заключается в том, что она не решает проблему происхождения жизни, а просто переносит ее в другое место (Приложение А на стр. 40). Хотя в самом строгом смысле это может быть верно, важность знания о том, зародилась жизнь или нет, или могла ли она это сделать de novo11 в самой крошечной космической среде (здесь, на Земле), когда речь идет о космосе в целом, является научным вопросом первостепенной важности и именно таким, ответ на который необходимо найти. Космическая теория жизни, которая расширяет интерактивную биосферу всего живого, чтобы охватить включающий все обитаемые ниши во Вселенной космологический объем, имеет глубокие ответвления в рамках самой эволюционной биологии. Некоторые из этих ответвлений появятся в других разделах этой статьи, а более подробное обсуждение точек зрения по поводу происхождения во Вселенной жизни per se12 можно найти в дополнительной информации (Приложение А на стр. 40).
По всей видимости, превращение ансамбля правильно выбранных биологических мономеров (например, аминокислот, нуклеотидов) в примитивную живую клетку, способную к дальнейшей эволюции, т.е. событие, которое не могло произойти в пределах временных рамок Земли, кроме как, на наш взгляд, чудесным образом (Hoyle and Wickramasinghe, 1981, 1982, 2000), требует преодоления информационного барьера суперастрономических масштабов (Приложение А на стр. 40). Все лабораторные эксперименты, пытающиеся смоделировать подобное событие, до сих пор кончались удручающими провалами (Deamer, 2011; Walker and Wickramasinghe, 2015). Поэтому представляется разумным обратиться к самому большому из доступных “месту действия” с точки зрения пространства и времени. Тем самым, космологическое происхождение жизни кажется нам правдоподобным и в высшей степени вероятным, а различные идеи, имеющие отношение к этому вопросу, были глубоко исследованы Хойлом, Викрамасингхе (Hoyle and Wickramasinghe, 1979, 1981, 1982, 2000) и Гибсоном (Gibson et al.,2011).
Благодаря использованию новых усовершенствованных инструментов и телескопов, ученые продолжают находить межзвездные органические молекулы все более нарастающей по сложности структуры. Для выявления присутствия таких молекул используются инфракрасные, микроволновые и радионаблюдения, и, вероятно, только
11 Заново (Прим.пер.)
12 Как таковой (Прим.пер.)
ограниченные доступные методы могут помешать продолжить этот список важных открытий. Уже давно было обнаружено, что первый средне-инфракрасный спектр инфракрасного источника GC-IRS7 в галактическом центре очень похож на спектр, который ранее был описан для частично разложившейся (лиофилизированной) бактерии (Рис.1), и эта поразительно точная корреляция между лабораторными данными и астрофизическими наблюдениями была обоснованно истолкована Хойлом и Викрамасингхе как убедительное доказательство того, что жизнь является космическим явлением (Hoyle et al., 1982, 1984).
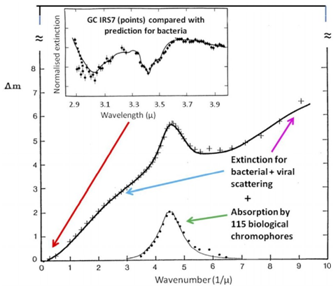
Рис.1 Сочетание свойств поглощения и рассеяния межзвездной пыли от инфракрасной до вакуумной областей ультрафиолетового спектра. [Ось абсцисс
- Волновое число (1/мкм); Ось ординат - Δm; Красная стрелка - длина волны (мкм) - прим.пер. рис.1] Самая низкая кривая - [Зеленая стрелка: Поглощение 115 биологических хромофоров - прим.пер. рис.1] - это прогнозируемое поведение объединения биологических ароматических молекул около 2175Å по сравнению со средними характеристиками межзвездной пыли; Средняя кривая - [Голубая и фиолетовая стрелки: поглощение бактериального+вирусного рассеяния - прим.пер. рис.1] - поведение при полном поглощении (поглощение + рассеяние) объединения бактериальной и вирусной пыли по сравнению с астрономическими
значениями данных для межзвездной пыли; Верхняя кривая - [Нормализованное поглощение. Значения GC IRS7 значения в сравнении с прогнозированием для бактерий - прим.пер. рис.1] представляет собой измеренное поглощение обезвоженных бактерий. Точки представляют собой астрономические наблюдения Д.Т.Викрамасингхе и Д.А.Аллена за источником галактического центра GC-IRS7 (1986). На основе опубликованных данных NCW. Подробнее см. ссылки (Wickramasinghe, 2015а, 2015b; Hoyle et al.,1982, 1984).
Доказательства, накопленные в последующие три десятилетия, только укрепили это представление: смесь полубитуминозного угля и высушенных бактерий E-coli дала такое же соответствие ИК-спектральным характеристикам GC-IRS7 на длине 3,2-3,8 мкм, как показано на Рис.2 (Coulson and Wickramasinghe, 2000), предполагая, что процесс бактериальной деградации приводит к образованию межзвездного угля.
Стандартным опровержением этой биологической интерпретации спектроскопических данных было утверждение, что взвешенное надлежащим образом объединение органических функциональных групп (полученных абиотически), которое точно соответствует такому биологическому спектру, может быть воспроизведено в биохимической или биофизической лабораториях. Но условия получения такой безошибочно точно настроенной смеси, которые можно было бы использовать повсеместно, никогда не исследовались и не публиковались теми, кто руководствовался такой точкой зрения. (Хотя скептическое научное отношение к таким вопросам имеет важное значение, мы, тем не менее, вновь подчеркиваем, что определение химических и
физических свойств внеземных объектов и сущностей на просторах Солнечной системы и всей Вселенной на основе корреляционных спектроскопических данных через телескопы в земных лабораториях было хлебом насущным для астрофизической дедуктивной науки, по крайней мере, со времен Галилея и эпохи Возрождения, как например, показывают имеющиеся данные на рис.1-3).
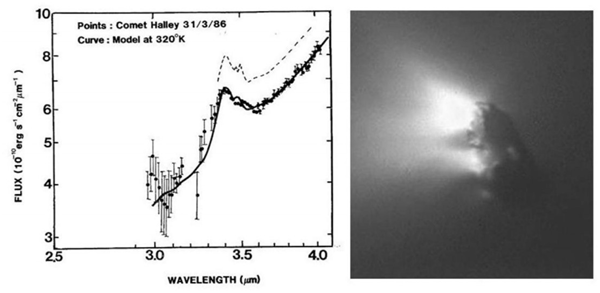 |
Рис.2 Эмиссия пылевой комой кометы Галлея 31 марта 1986 г. (точки) по сравнению с нормализованными потоками для высушенной кишечной палочки при температуре эмиссии 320К. Сплошная кривая - необлученные бактерии; пунктирная кривая - бактерии, облученные рентгеновскими лучами (Allen and Wickramasinghe, 1981; Wickramasinghe and Allen, 1986). График слева взят из опубликованных данных N.C.W. Справа, изображение ядра кометы Джотто (снимок предоставлен Европейским космическим агентством).
Доказательные доводы в пользу того, что кометы обладают сложной органикой впервые были выдвинуты В.Ванисеком и Ч.Викрамасингхе (Vanýsek and Wickramasinghe, 1975). В 1986 г. присутствие органической пыли в кометах было подтверждено Д.Т. Викрамасингхе и Д.А. Алленом (D.T. Wickramasinghe and Allen,1986), рис.2. Впоследствии Хойл и Н.Ч.Викрамасингхе (Hoyle and N.C. Wickramasinghe,1986) отметили, что инфракрасный спектр кометы Галлея указывает на наличие в кометах подобного бактериям вещества (Рис.1), а также на то, что кометы, по-видимому, имеют смолистый поверхностный слой, образовавшийся в результате разложения биологического материала вблизи ближайшей орбитальной точки около Солнца, или перигелия.
Эти данные бросили серьезный вызов модели Уиппла “грязный снежок”13, которая была господствующей парадигмой того времени. Космические зонды кометы Галлея установили, что пыль с высоким содержанием углеродистых соединений и очень темная поверхность делают ее совершенно не похожей на лед или снег. Более позднее изучение нескольких комет с использованием широкого спектра космических технологий усилило
13 Ф.Л.Уиппл, американский астроном, предложивший модель ядра кометы как смеси льдов с вкрапленными частицами метеорного вещества (теория «грязного снежка»). (Прим.пер.)
доводы в пользу наличия в кометах микробной жизни (и в углеродистых остатках хондрита), но консерваторы из числа астрономов и специалистов по метеоритам не готовы с легкостью это признать. Предполагается, что короткопериодические кометы Солнечной системы, а также долгопериодические кометы, источником которых является Облако Оорта и соседние звездные системы, являются носителями и распространителями микробной жизни в галактическом или даже космологическом масштабе (Hoyle and Wickramasinghe, 1981, 1993). Из этой модели следует, что межзвездная пыль должна включать в себя часть материала, которая представляет собой биологические детриты.
 Рис.3 Струи органических молекул и молекулярного кислорода, исходящие из кометы 67P/C-G (Предоставлено Европейским космическим агентством). Соотношение P/C газа, проанализированного орбитальным аппаратом Rosetta, выявило значения 1%, который соответствует условию разложения бактерий (Capaccionne et al., 2015, Bieler et.al.,2015, Altwegg et al., 2016).
Рис.3 Струи органических молекул и молекулярного кислорода, исходящие из кометы 67P/C-G (Предоставлено Европейским космическим агентством). Соотношение P/C газа, проанализированного орбитальным аппаратом Rosetta, выявило значения 1%, который соответствует условию разложения бактерий (Capaccionne et al., 2015, Bieler et.al.,2015, Altwegg et al., 2016).
С 1980 г. присутствие в межзвездных облаках сложных органических молекул, таких как полициклические ароматические углеводороды, не подлежит сомнению (Hoyle and Wickramasinghe, 1991, 2000). В дополнение к данным о инфракрасном диапазоне повсеместная полоса поглощения 2175Å в межзвездной пыли, хотя до сих пор «не идентифицирована», по-видимому, полностью соответствует либо биохимическим хромофорам (разрушение микробиологии), либо преобразованным радиацией микробным образованиям. Другой набор астрономических сведенией, указывающих на повсеместное присутствие микробиологии, - это диффузные полосы межзвездного поглощения в визуальных спектрах звезд, которые не поддавались идентификации более 8 десятилетий, но которые соответствуют свойствам порфиринов ((Hoyle and Wickramasinghe, 1991). Также следует отметить, что за последние 15 лет список соответствия, показанный на рис.1, был значительно расширен и теперь включает самые далекие галактики (Wickramasinghe, 2015a). Это означает, что случайное совпадение биохимических спектров с астрономией должно (по мнению критиков и скептиков) простираться до самого края наблюдаемой Вселенной (красное смещение z=4).
Посадочный модуль Philae миссии Rosetta недавно предоставил нам новую информацию о комете 67P/C-G (Capaccionne et al., 2015; Walker and Wickramasinghe, 2015; Wickramasinghe et al., 2015). Струи пара H2O и органических веществ, исходящие из трещин и отверстий в черной коре (рис. 3), вероятно, соответствуют биологической активности, происходящей в подповерхностных водоемах (Wickramasinghe et al., 1996, 2009). Последний отчет об O2 вместе с доказательствами наличия воды и органических веществ, на наш взгляд, является
еще одним убедительным аргументом в пользу продолжающейся биологической активности (Bieler et al., 2015). Такая смесь газов не может быть получена в термодинамических условиях, поскольку органические вещества легко разрушаются в окисляющей среде. Был предложен вариант замерзания исходной смеси соединений, включая O2, не в термохимическом равновесии, но нет никаких доказательств, чтобы подтвердить такое утверждение. С другой стороны, отток органических веществ O2/H2O из кометы можно изящно объяснить условиями подповерхностной микробиологии. Фотосинтезирующие микроорганизмы, функционирующие в перигелии у поверхности при низких уровнях освещенности, могут производить O2 вместе с органическими веществами. Многие виды ферментирующих бактерий могут также производить этанол из сахаров, поэтому недавнее открытие, что комета Лавджой (Lovejoy) выделяет этиловый спирт в количестве, эквивалентном 500 бутылок вина в секунду, вполне может быть признаком того, что такой микробный процесс работает (Biver et al., 2015).
Научная история вопроса о жизни на Марсе не только представляет отдельный интерес сама по себе, но и весьма значима в контексте космической биологии (Hoyle and Wickramasinghe, 1997, Wickramasinghe, 2015a). Ранние положительные результаты Гилберта Левина и Патрисии Страат (Gilbert Levin and Patricia Straat,1976) о сохранившейся микробной жизни, обнаруженной на поверхности Марса в эксперименте Viking Labeled Release (LR) 1976 года, так и не были должным образом опровергнуты (Levin and Straat, 2016), при этом Левин и его коллеги всесторонне рассматривали и учитывали всевозможные комментарии и критические замечания (Levin, 2007, 2013, 2015; Bianciardi et al., 2012). Несомненно, возможности для более качественного научного поиска жизненных миссий на Марсе с участием Гилберта Левина улучшились в значительной мере (пресс-релиз Букингемского университета, 23 июля 2016 г.). Новаторские исследования 1976 года подтвердились новыми результатами Раффа и Фармера (Ruff and Farmer, 2016), полученными с марсохода Mars Spirit, который обнаружил на Марсе отложения диоксида кремния, содержащие элементы, напоминающие биосигнатуры горячих источников в Эль-Тайоне в Чили. Рафф и Фармер заключили, что “хотя целиком абиотические процессы для структур марсианского диоксида кремния не исключены, они удовлетворяют априорному определению потенциальных биосигнатур”. Поэтому вполне уместно, что тот обширный набор данных, который относится к вероятности существования жизни на Марсе, был в полной мере оценен, а открытия Левина и Страата, которые были ими сделаны четыре десятилетия назад, теперь пришлись к месту, и оба ученых получили должное признание за свои исследования.
- Микробный материал в стратосфере и в метеоритах
Данные кометных исследований по-прежнему подкрепляются извлечением микробного материала в стратосфере (в условиях, когда апвеллинг14 земного загрязнения может быть логично исключен). В стратосфере, на высоте диапазоном 30-41 км, продолжают обнаруживать биологические структуры: от жизнеспособных, но не культивируемых микробов, до необъяснимых скоплений микроскопических биологических образований (Wainwright et al., 2014, 2015а, 2015b). Структуры состоят из углерода и азота, обладают двусторонней симметрией и организмоподобной морфологией. Свидетельства были интерпретированы так, чтобы показать, что они согласуются с приемлемым выводом о том, что эти похожие на микроорганизмы сущности прибывают на Землю из космоса и, возможно, переносятся небольшими кометами (Frank and Sigwarth, 2001). Была доказана вероятность существования биологических материалов, спускающихся через атмосферу Земли, в метеороидах микронного размера (Coulson and Wickramasinghe, 2003) и в ледяных кометных метеорах радиусом ~1 м. (Coulson et al., 2014). Стали хорошо известны ранние доказательства наличия окаменелых микроорганизмов внутри углеродистых метеоритов, и теперь эти данные дают уверенный отпор скептицизму по поводу загрязнения земли (Pflug and Heinz, 1997; Hoover, 2005,2011; Miyake et.al.,2010; Wickramasinghe, 2015a). Еще одним убедительным доказательством панспермии служит последнее обнаружение микробных окаменелостей в метеоритах, упавших в Шри-Ланке в 2012 году, за которым последовало недвусмысленное определение внеземного происхождения их пород (на основе данных изотопов кислорода) (Wallis et al., 2013). Другой похожий феномен касается явления красных дождей, которые фиксировались на протяжении истории (McCafferty, 2008), и случились совсем недавно в Керале, Индии (Louis and Kumar, 2006) и в Шри-Ланке в 2012 году. Все имеющиеся доказательства указывают на краснопигментированные организмы, которые вряд ли имеют земное происхождение.
Несмотря на тщательные меры предосторожности и контроля, которые были предприняты для исследований, приведенных нами в этом разделе, общая тенденция заключается в том, чтобы отвергать подобные открытия, поскольку они противоречат господствующей парадигме и дискредитируют ее.
- Принципы вирусологии - вирусы как плотные информационно-насыщенные системы контроля
Прежде чем приступить к обсуждению новых данных об эволюции ретровирусов (Aiewsakun and Katzourakis, 2017), важно вспомнить основные процессы биологии вирусов и их modus operandi15 как генетических векторов внутри и между клетками.
14 Термин использован здесь по аналогии с: Апвеллинг - подъем глубинных вод океана к поверхности. Различают как минимум четыре типа апвеллинга: прибрежный; крупномасштабный; ветровой в открытом океане; связанный с вихрями; связанный с топографией. (Прим.пер.)
15 Принцип работы (от лат. “образ действия”). (Прим.пер.)
Все ДНК- и РНК-вирусы, заражающие бактериальные или эукариотические клетки, представляют собой жестко упорядоченные и плотные информационные системы. Все известные клеточные системы несут интегрированную генетическую информацию вируса или ее фрагменты и вследствие этого являются потенциальными целями вирусной атаки и заражения. Эти аксиомы не противоречат ни одному из принципов ни в одном современном учебнике. Действительно, с тех пор, как Феликс д'Эрелль (Felix d’Herrelle) и Фредерик Туорт (Frederick Twort) открыли «фильтруемые» агенты в 1915-17 годах, которые могут инфицировать и убивать бактериальные клетки, у нас появилось почти что полное понимание существенных принципов вирусологии.
Чтобы продолжить, кратко суммируем то, что мы знаем:
- Накопленный на протяжении десятилетий экспериментальный опыт с системами трансформации и трансфекции позволяет сделать вывод о том, что любая вирусная частица или агент нуклеиновой кислоты (вироид) в принципе потенциально может проникнуть в любую клетку.
- Репликация внутри клетки - это другое дело, поскольку входящая информационно-насыщенная молекулярная система должна одновременно образовывать гель, сцепляться и интегрироваться в сложную биохимическую и генетическую структуру клетки-хозяина (Рис.4).
- Но сложная репликация может привести к резкой выработке вирионов и гибели клетки-хозяина (см. Цитопатические вирусы). Такие явления происходят из-за того, что и непосредственный механизм врожденного иммунитета клетки, а затем и замедленный ответ приобретенного иммунитета организма, не в состоянии остановить вирусную инфекцию. Резкая выработка вирионов происходит реже, чем более умеренная репликация и внеклеточный экспорт вирионов, что способствует росту как вируса, так и клетки-хозяина (или даже интеграции вирусной ДНК/РНК в геном клетки-хозяина в качестве латентного вируса). Таким образом, таковы три основных итога первой стадии взаимоотношений паразит - хозяин (Рис. 4).
- Обычно инфекция типа резкого цитопатического роста (ведущая к быстрой гибели хозяина) встречается редко, потому что большинство циркулирующих (и развивающихся) Земных вирусов, установили правила ведения боевых действий “паразит - хозяин” (как в своих встроенных, так и в приобретенных средствах защиты).
- Таким образом, острые цитопатические инфекции, характерные для неожиданно быстро возникающих эпидемий и пандемий, вероятно, при первом проникновении имеют основания, на которые обычные ограничения на взаимоотношения по принципу “паразит - хозяин” не накладываются (см. ниже).
- Следовательно, круг хозяев всех вирусов среди растений, животных и бактерий определяется не проникновением вириона в клеточное микроокружение, а тем, может ли он продуктивно реплицироваться или интегрироваться, а затем экспрессироваться (например, ретровирус). На практике это обычно проявляется в виде заметного вирусного заболевания с характерными не опасными для жизни первыми симптомами.
- Таким образом, вирусы представляют собой информационно-насыщенные полинуклеотидные макромолекулы размером ≥3000 п.н.16 (SanJuan, 2010, 2012;Sanjuan and Domingo-Calap, 2016). В восприимчивых клетках-хозяевах они в некотором смысле представляют собой сжатые регуляторные схемы, согласованные с основными генами регуляторов транскрипции и узловыми биохимическими путями, критическими для роста и жизнеспособности клетки.
- В таком случае мы должны аргументированно рассматривать вирусы как одну из самых информационно-насыщенных природных систем в обозримой Вселенной (Рис.4). Размер вирусов делает их очень маленькими мишенями, что сводит к минимуму вероятность гибели от импульсного нагрева или ионизирующего излучения (Hoyle and Wickramasinghe, 1979, напр., Глава 1). Благодаря нанометровым размерам, их, вероятно, могут легко транспортировать и распылять частицы пыли микрометрового размера и другие защитные физические матрицы аналогичного размера. В таком случае они представляют собой генетические векторы размером с наночастицы, которые содержат всю существенную информацию, необходимую для захвата и управления физиологией любой заданной клетки-мишени, с которой они сплелись изнутри. Их репликативный рост означает, что они производятся и существуют в огромных количествах в космических масштабах; вследствие этого, они (и в меньшей количественной степени их клеточные резервуары) могут нести огромные потери из-за инактивации, но в то же время оставляя следы из миллионов выживших частиц, потенциально все еще вирулентных. Таким образом, вирус представляет собой тип сжатого модуля, который связан с самой способностью клетки расти и делиться, чтобы производить клетки-потомки и, таким образом, эволюционировать.
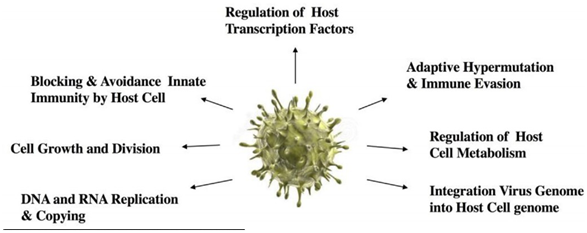 |
Рис.4 Типичные основные функции вирусов17 находятся в соответствии со всеми существенными аспектами роста собственного воспроизводства и более широкого клеточного строения. Изображение вируса: https://www.dreamstime.com/royalty-free-stock-photo-h1n1-viruses-image11898035
16 П.н. - пара нукдеотидов, или нуклеотидных оснований. (Прим.пер.)
17 Слева направо по часовой стрелке: Репликация и копирование ДНК и РНК - Рост и деление клеток - Блокирование и отказ от врожденного иммунитета клеткой-хозяином - Регуляция факторов транскрипции клетки-хозяина - Адаптивная гипермутация и уклонение от иммунитета - Регуляция метаболизма
клетки-хозяина - Интеграция генома вируса в геном клетки-хозяина (Прим.пер.)
Эти ключевые моменты основаны на результатах важных экспериментов Санхуана и его помощников над РНК-вирусами (Sanjuan, 2010, 2012; SanJuan et al., 2004, 2010; Combe et al., 2015). Предполагается, что точно отлаженные информационно-насыщенные вирусные геномы могут быть подвержены случайным мутациям, снижающим скорость их репликации и вирулентности. Использование структурно-направленного мутагенеза для получения случайных мутаций в вирусных геномах (SanJuan et al., 2004; Sanjuan, 2010) до 40% таких мутировавших нуклеотидов приводит к гарантированному летальному исходу. Это только приблизительная оценка, поскольку замены, приводящие к замещению аминокислот (несинонимичные мутации), - это не единственная мутационная угроза, с которой может столкнуться вирус. Тихие замены в области 3’-конца цепочки “неоднозначных пар” в кодоне могут иметь функциональные последствия, влияющие на репликацию и вирулентную эффективность, которые вызваны котрансляционными паузами / задержками при выходе полипептида из рибосомы. Эти эффекты приводят к аномалиям кинетики сворачивания из-за вариаций в составе объединения прекурсоров нуклеозидтрифосфата (Buhr et al., 2016). Хорошо сохраненные структуры РНК в геноме РНК HCV18 при манипулировании синонимичными центрами сегментов в области 3’-конца могут изменять способность вируса к репликации и, следовательно, инфицированию (Pirakitikulr et al., 2016).
Изощренность вирусной вирулентности и ее modus operandi по распространению клеток по клеткам на этом не заканчивается. Чтобы обеспечить максимальную комплементацию генов, дефектные вирусные геномы, несущие стоп-кодоны (Aaskov et al., 2006), можно размножать почти бесконечно (Combe et al., 2015) как «кластеры вирионов» из смесей инфекционных и поврежденных геномов со значительным количеством вновь созданных вирусных частиц, заключенных в везикулы защитных мембран, тип многокомпонентной наночастицы. Тогда это образует фактическую инфекционную дозу, а не просто один экспортированный вирион, попадающий в соседнюю клетку-мишень, чтобы вызвать эффективное заражение, как обычно считается (Combre et al.,2015, Chen et al.,2015).
Однако необходимо еще раз указать, что большинство Земных вирусов уже развили установившиеся молекулярно-интерактивные отношения паразит - хозяин. Многие из них сами по себе являются доброкачественными и нецитолитическими, поэтому многие начальные инфекции обычно протекают тихо и бессимптомно. Врачи и клиницисты теперь понимают, что основной проблемой для здоровья часто является непреднамеренное повреждение тканей, вызванное ответом в виде воспалительных реакций со стороны приобретенного иммунитета пациента, когда ткани запущены до состояний хронической инфекции, как например при вирусе HCV гепатита (у тех пациентов, которым не удается быстро и естественным путем избавиться от первоначальной инфекции).
Кажется, нет конца изобретательным стратегиям, которые вирусы используют для заражения и захвата своих клеток-мишеней, будь то бактериальные (нормальные прокариоты, экстремофильные архебактерии) или эукариотические клетки многоклеточного животного. Если мы просто рассмотрим недавний образец РНК-вирома беспозвоночных носителей, сложная величина которого - с обменом и совместным использованием последовательностей вирусных генов - просто поражает своими
18 Вирус гепатита С (Прим.пер.)
масштабами (Shi et al., 2016). У более широко изученных позвоночных, у которых случились аналогичный обмен и замещение генов, уклонение как от врожденного, так и от приобретенного иммунного ответа является ключевыми аспектами жизненного цикла вируса. Недавно было замечено, что одна из возможных причин, по которой ретровирус ВИЧ не может контролироваться обычными иммунологическими вакцинами (вырабатывающими нейтрализующие антитела до заражения вариантом ВИЧ-1), заключается в том, что основным клеточным центром провирусной интеграции может быть не Т-лимфоцит или макрофаг/дендритная клетка, как обычно предполагают, но, более вероятно, В-лимфоцит (Steele and Dawkins, 2016), клеточный дом механизма соматической гипермутации (Franklin et al., 2004; Steele, 2016b; Steele and Lindley, 2017). Таким образом, было высказано разумное предположение, что ВИЧ, используя механизм адаптивной соматической гипермутации системы антител, всегда будет на один мутационный шаг впереди собственного приобретенного иммунного ответа пациента (Steele and Dawkins, 2016). С тех пор, как это предположение было озвучено, оно не было никак публично оспорено основными критиками в областях иммунологии и вирусологии, и остается концептуальным вариантом для лучшего понимания modus operandi ВИЧ и связанных с ним ретровирусов.
- Критика особенностей хозяина-носителя вирусов и его выживаемость в космических условиях
Критика, направленная против концепции вирусов, вызывающих болезни и вносящих свой вклад в эволюцию жизни, сводится к одному риторическому вопросу: как вирус из космоса может знать до своего прибытия сюда ряд организмов, которые будут ему доступны и с которыми он сможет взаимодействовать? В предыдущем разделе мы проработали правильный ответ на этот вопрос: возникающие в космических условиях вирусы и эволюция на Земле - неразрывно связаны. Особенности хозяина-носителя вирусов сохраняются только для довольно узкого ряда видов, определяемых их эволюционной историей. Так, вирус гриппа А можно культивировать в куриных яйцах, что подтверждает особенности его хозяина на протяжении 50 миллионов лет эволюционной истории (и известно, что он заражает многие виды позвоночных и млекопитающих). Опять-таки, вирус Эбола поражает класс приматов.
Критика того, что бактерии и/или вирусы неспособны выжить в суровых условиях космоса, определенно не подтверждается всеми данными, накопленными за последние три десятилетия. Бактерии и вирусы, внедренные в песчинки, углеродистый материал или лед, эффективно защищены от радиационного повреждения и могут оставаться полностью жизнеспособными в течение миллионов лет в космических условиях. Микроорганизмы, включая вирионы, глубоко замороженные внутри кометных тел, могут оставаться жизнеспособными бесконечно, и, конечно, в космологических масштабах времени. Недавние космические эксперименты, в том числе проводимые на борту Международной космической станции, показали поразительную способность бактерий и вирусов к выживанию.
Вследствие этого, К.С.Тиль19 и ее коллеги (Нiel et al., 2014) недавно провели эксперимент, в котором плазмидная ДНК была помещена на внешнюю поверхность ракеты-зонда TEXUS-49, которая была выпущена через атмосферу в космос и позднее снова вошла в атмосферу. Условия, которым подверглась ДНК, точно имитировали то, что на самом деле происходило бы при высокоскоростном проникновении вирусов, прикрепленных к метеорной/кометной пыли. Результаты, полученные К.С.Тиль и ее коллегами (Нiel et al., 2014) показали, что значительная часть ДНК осталась жизнеспособной и вирулентной, а это явный признак того, что внеземные вирусы действительно могли прибыть на Землю в жизнеспособной форме.
Было обнаружено, что микроорганизм D.audaxviator20, обнаруженный на глубине 2,8 км на золотом руднике в Южной Африке, получает свою энергию от радиолиза, вызванного частицами, испускаемыми при распаде U, Th и K21 (Chivian et al., 2008) . Такие бактерии, как D.audaxviator, выживут не только в межзвездных транзитах, но и будут процветать за счет энергии, полученной от галактических космических лучей, которые достигают межзвездных или межпланетных замороженных тел.
- Модель индукции ретровирусов
Кроме того, теперь мы гораздо лучше понимаем четкие недарвиновские и неменделевские механизмы эволюционного наследования, формирующих, в частности, как иммунную, так и центральную нервную систему (например, работы Ламарка о ретровирусах и РНК/RТ22) (Steele, 1979; Gorczynski and Steele, 1980,1981;Steele et al., 1984; Steele et al., 1998; Steele, 2016a), а также ретровирусные/ретроэлементные драйверы сегментарных дупликаций и структуры геномного блока предковых гаплотипов и связанных с ними явлений недарвиновского наследования, имеющих медицинское значение (Dawkins et al., 1999; Dawkins, 2015; Steele, 2014, 2015; Steele and Lloyd, 2015). В целом, в разных источниках обсуждается множество механизмов адаптивного ламарковского наследования (Campbell, Perkins, 1988; Jablonka, Lamb, 1995; Lindley, 2010; Liu, 2007; Noble, 2013; Mattick, 2012; Liu and Li, 2016a, 2016b), некоторые из которых связаны с подвижными лимфоцитами, доставляющими эндогенные ретровирусы и соматические гены к клеткам зародышевой линии (Rothenfluh, 1995), или рассматривались другие типы механизмов передачи от сомы к зародышевой линии с участием везикул или экзосом (Spadafora, 2008; Cossetti et al., 2014; Devanapally et al., 2015; Sharma et al., 2015). Таким образом, можно выделить и рационально объяснить четкую причинно-следственную связь новых вирусов, прибывающих из космоса и потенциально способствующих эволюции на Земле (Wickramasinghe and Steele, 2016). Действительно, транспозиция ретроэлементов LINE (и совместная мобильность элементов Alu-повторов, Приложение B на стр. 44) является
19 Имя автора, о котором идет речь, - Cora S. Thiel. В оригинальном тексте статьи произошла опечатка, вследствие чего ошибочно указывается фамилия Hiel, включая адреса ссылок. (Прим.пер.)
20 Candidatus Desulforudis audaxviator (лат.) — уникальный вид экстремофильных анаэробных бактерий, живущих на глубинах от 1,5 км до 3 км ниже поверхности земли в подземных водах, способных существовать обособленно от каких-либо других живых организмов. (Прим.пер.)
21 Хим. элементы U (Уран), Th (Торий) и К (Калий) (Прим.пер.)
22 От англ. Reverse Transcriptase - Обратная Транскрипция (Прим.пер.)
нормальной частью геномной перестройки во время специфической фиксации нейрона, во многом подобной перестройке V➝DJ в специфической фиксации B- и Т-лимфоцитов в иммунной системе (Erwin et al., 2016).
Что касается ВИЧ и ретровирусной эволюции в целом, а именно, геномных дупликационных процессов, генерирующих полиморфную блочную (предковую) гаплотипическую структуру человеческого генома, основные концепции можно проследить вплоть до известной сейчас «Модели индукции ретровирусов» (Dawkins et al., 1999; Steele, 2014; Steele, 2015, стр.95). Таким образом, когда ретровирус заражает человеческую клетку, запускаются все способы мутагенных процессов, включая: индуцированные AID/APOBEC-дезаминазой явления C-to-U, приводящие к мутациям C-to-T, места Abasic и разрывы оцДНК, а также явления редактирования РНК A-to-I, индуцированного ADAR-дезаминазой. В настоящее время оба эти дезаминирования ДНК и РНК идентифицированы как целевые соматические мутации (ЦСМ), связанные с цепью и условиями кодона, в геноме рака человека (Lindley, 2013; Lindley and Steele, 2013; Lindley et al., 2016; Steele and Lindley, 2017). Кроме того, у нас есть мутагенная мобильность LINE/Alu-ретроэлемента (Harris and Liddament, 2004; Chiu et al., 2006; Muotri et al., 2007; Doria et al., 2009; Refsland and Harris, 2013; Jones et al., 2013). Итак, как уже обсуждалось, ретромобильность LINE/Alu теперь рассматривается как нормальный этап развития специфических синаптических нейронов мозга (Erwin et al., 2016). Редактирование РНК (A-to-I), нацеленное на нейронные инвертированные Alu элементы (в интронах, создавая альтернативные сплайсированные изоформы), и является признанным процессом синаптической нейронной диверсификации в головном мозге (Paz-Yaacov et al., 2010). Таким образом, ретровирусы и другие вирусы, которые, предположительно, высвобождаются из следов кометных обломков, потенциально могут добавлять новые последовательности ДНК в земные геномы и вызывать дальнейшие мутагенные изменения в геномах соматической и зародышевой линий (Приложение B на стр. 44). Действительно, Фрэнк Райан назвал управляемую вирусами земную эволюцию, используя соответствующий крылатый термин «виролюция», и эта концепция была поддержана и расширена Оливером и Грином (Oliver and Greene, 2012) в Гипотезе направленности переносимых элементов (Приложение B на стр. 44). Д.Йон и его коллеги (Yohn et al. (2005)) заявили, что их данные согласуются с ретровирусной инфекцией, которая 3-4 миллиона лет назад самостоятельно и параллельно поразила геномы шимпанзе и горилл без горизонтального заражения. Недавно Диль и его соавторы (Diehl et al.,2016) доказали, что около 33-15 миллионов лет назад определенная группа эндогенных ретровирусов (ERV-Fc) каким-то образом стала распространяться по всему миру среди многих видов млекопитающих. Одновременно были высказаны мнения, что более позднее эволюционное развитие гоминидов, включая усиление когнитивных способностей, также могло иметь вирусное происхождение (Villareal, 2004). Предполагается вероятное проникновение внешнего вируса в плаценту у предков всех млекопитающих, включая человека, около 150 миллионов лет назад (Katzourakis, 2013).
- Эволюция усложнения интеллектуальной комплексности
Недавно при секвенировании генов и транскриптомов головоногих моллюсков были получены предполагаемые доказательства роли внеземных вирусов в их влиянии на земную эволюцию. Уровень строения генома осьминога демонстрирует ошеломляюще сложную структуру - он содержит на 33000 кодирующих ген белков больше, чем геном Homo sapiens (Albertin et al., 2015). Осьминог принадлежит к подклассу моллюсков (головоногих моллюсков), эволюционная история которых насчитывает более 500 миллионов лет, хотя филогенетика головоногих моллюсков очень непоследовательна и сбивает с толку (см.:Carlini et al.,2000; Strugnell et al., 2005, 2006, 2007; Bergmann et al., 2006). Головоногие моллюски также очень разнообразны: сложные по поведению колеоиды23 (кальмары, каракатицы и осьминоги) предположительно появились в рамках чисто земной эволюционной модели из более примитивных наутилоидов. Однако генетическое отличие осьминога от его предкового подкласса колеоидов очень велико, и его некоторые специфические черты больше схожи с характеристиками многих родов и видов, отмеченных в паттернах прерывистого равновесия Элдриджа-Гулда (ниже).
 |
Рис.5 Эволюция от кальмара до осьминога согласуется с набором генов, внедренных внеземными вирусами. Обсуждается альтернативный внеземной сценарий, согласно которому популяция криоконсервированных эмбрионов осьминога мягко приземлилась en mass24 из космоса 275 миллионов лет назад. Изображение кальмара - Grimalditeuthis bonplandi. Изобр. Wikimedia Commons. Автор: Jeanne Le Roux & L. Joubin URL (http://www.archive.org/stream/rsultatsdescam17albe#page/n165/mode/2up0). Изображение вируса из: https://www.dreamstime.com/royalty-free-stock-photo-h1n1-viruses-image11898035.
Его большой мозг и сложная нервная система, похожие на объектив глаза, гибкое тело, мгновенная маскировка за счет способности переключать цвет и форму - это лишь некоторые из поразительных особенностей, внезапно появляющихся на эволюционной сцене. Трансформирующиеся гены, наследуемые от древнего наутилуса (например, Nautilus pompilius) к обыкновенной каракатице (Sepia officinalis), кальмару (Loligo vulgaris) и обычному осьминогу (Octopus vulgaris, рис.5), нелегко найти ни в одной из ранее существовавших форм жизни. Поэтому можно предположить, что они заимствованы из
23 Колеоиды, или Двужаберные - подкласс головоногих моллюсков (Прим.пер.)
24 От лат. - Всей массой (Прим.пер)
далекого «будущего» с точки зрения земной эволюции или, что более реалистично, из космоса в целом.
Такие объяснения происхождения жизни через внеземные источники, конечно, противоречат существующей доминирующей парадигме. Однако с этим выводом согласуются недавние, основанные на редактировании РНК всего транскриптома данные о механизмах диверсификации соматической РНК у сложных в поведении головоногих моллюсков, таких как осьминог. Эти данные демонстрируют обширные эволюционные консервативные сайты редактирования мРНК от аденозина к инозину (A-to-I) почти в каждом гене, кодирующем белок, у поведенчески сложных колеоидных головоногих моллюсков (в частности, осьминога), но не у наутилуса (Liscovitch-Brauer et al., 2017). По сравнению с наутилусом и другими беспозвоночными и позвоночными животными, это огромное качественное различие в редактировании мРНК A-to-I, кодирующих белок головоногих моллюсков, просто поразительно. Таким образом, при секвенировании всего транскриптома было обнаружено, что только 1-3% мРНК дрозофилы и человека, кодирующие белок, несут сайты перекодирования A-to-I; и есть всего около 25 сообщений человеческих мРНК, которые содержат консервативную структуру перекодирования A-to-I среди млекопитающих. У линии дрозофил имеется около 65 консервативных A-соединений в генах, кодирующих белки, и лишь несколько из них, идентифицированных у C.elegans25, подтверждают гипотезу о том, что перекодирование редактирования РНК A-to-I в основном либо нейтрально, либо вредно, либо редко адаптивно (см.:Liscovitch-Brauer et al., 2017). Однако для кальмаров и особенно осьминогов это является нормой, поскольку почти каждый ген, кодирующий белок, имеет эволюционную консервативную изоформу сайта редактирования мРНК A-to-I, что приводит к несинонимичным аминокислотным изменениям (Liscovitch-Brauer et al., 2017). Этот виртуальный качественный скачок в молекулярно-генетической стратегии в предполагаемой плавной и постепенной эволюционной линии - тип внезапного “большого скачка вперед”. Если все новые гены, экспрессируемые в линиях кальмаров/осьминогов, не возникли в результате простых мутаций существующих генов либо у кальмаров, либо у других организмов, разделяющих ту же среду обитания, то определенно не существует способа, с помощью которого этот большой качественный переход в мРНК A-to-I редактирование можно объяснить обычными неодарвинистскими процессами, даже если разрешен горизонтальный перенос генов. На наш взгляд, одно из убедительных объяснений состоит в том, что новые гены, скорее всего, были импортированы на Землю из внеземных источников - наиболее вероятно в качестве уже целостной группы функционирующих генов внутри (скажем) замороженных и защищенных матрицей оплодотворенных яйцеклеток осьминога.
Таким образом, не следует сбрасывать со счетов вероятность того (ниже), что замороженные яйцеклетки кальмаров и/или осьминогов прибыли в ледяных болидах несколько сотен миллионов лет назад. По крайней мере, это могло бы быть лаконичным объяснением со стороны космоса, как осьминоги внезапно появились на Земле около 270 миллионов лет назад. Действительно, этот принцип применим к неожиданному возникновению в летописи окаменелостей практически всех основных форм жизни, охваченных пророческой концепцией «прерывистого равновесия», выдвинутой
25 Свободноживущая Нематода (круглый червь) (Прим.пер.)
Элдриджем и Гулдом в начале 1970-х годов (1972,1977); и см. Умозрительную схему на рис. 6. Поэтому схожие живые особенности, подобные “как если бы гены произошли от некоего типа предсуществования” (Hoyle and Wickramasinghe, 1981), при внимательном рассмотрении применимы ко многим другим биологическим объединениям. Один малоизвестный, но убедительный пример - это реакция и устойчивость глазных структур плодовой мухи дрозофилы к обычно смертельно опасному ультрафиолетовому излучению в 2537Å, учитывая, что эта длина волны не проникает через озоновый слой и, таким образом, не является очевидным дарвиновским селективным фактором на поверхности Земли (Lutz and Grisewood, 1934 и см.Hoyle and Wickramasinghe,1981, стр. 12-13). Многие из этих “неземных” свойств организмов могут быть логично объяснены, если мы признаем расширенную космическую биосферу, на которую указывают современные астрономические исследования и уже обсужденные открытия экзопланет. Среднее расстояние между пригодными для жизни планетами в нашей галактике сейчас измеряется в световых годах - обычно 5 световых лет (Wickramasinghe et al., 2012), - поэтому обмен вирионом/геном кажется неизбежным на таких коротких космических расстояниях. Многие особенности биологии, которые не оптимизированы для местных условий на Земле, могут быть легко объяснены в этой более широкой перспективе.
Исходя из того, что сложные наборы новых генов у осьминогов могли возникнуть не только в результате горизонтального переноса генов, простых случайных мутаций существующих генов или простых дублирующих расширений, можно логично предположить, учитывая наши текущие знания о биологии комет и их осколков, что новые гены и их вирусные носители, скорее всего, пришли из космоса. Помимо этого ясно, что принятие такого предположения также требует, чтобы мы уменьшили значение сильно локализованной дарвиновской эволюции на Земле, чему, вероятно, будут сильно сопротивляться сторонники традиционной биологии. Конечно, не должно вызывать беспокойства то, что фокус нашего внимания на общих эволюционных молекулярных процессах теперь смещается в Космос и за пределы нашей непосредственной солнечной системы. Эта информация обеспечивает и позволяет изучать космические генофонды, а они способны управлять биологической эволюцией здесь, на Земле, и, осмелимся сказать, контролировать ее (посредством дарвиновских и недарвиновских механизмов адаптации). Главный эффект Земной дарвиновской эволюции состоит в том, чтобы воздействовать на эти новые, полученные из космоса гены и откорректировать их с помощью дополнительных механизмов обратной связи ламарковских генов соматической и зародышевой линии и механизмов перетасовки гаплотипов в соответствии с окружающей средой и организмом-реципиентом (Wickramasinghe and Steele, 2016). Более того, было выявлено, что вирусные следы прослеживаются в тканях человеческого мозга, и это, кажется, знаменует важные этапы, которые привели человека к его нынешнему положению (Villarreal, 2004; Ryan, 2009).
Последствия смены парадигмы
для взаимосвязей филогенетической последовательности
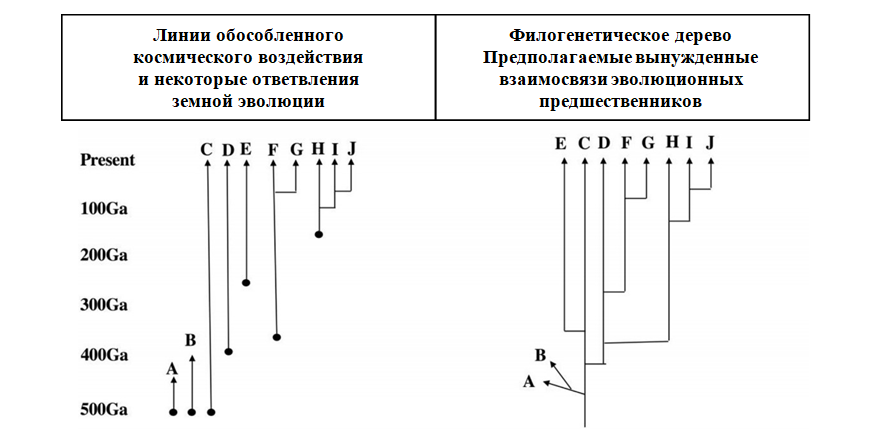
Рис.6 Умозрительный схема. Эволюция после воздействия из Космоса в сравнении с Эволюционными взаимосвязями в пределах Земли.26 Эволюционные закономерности для необходимого консервативного белка, такого как, например, гемоглобин (перенос O2) или цитохром C (перенос электрона). Десять различных вариантов белков - от A до J. Этот рисунок иллюстрирует концептуальные различия между схемами эволюции, вызванные проникновением разных видов организмов из космоса с последующей земной эволюцией (слева) по сравнению с эволюционными отношениями между теми же группами консервативных белков после после того, как филогенетическое дерево будет вынуждено использовать данные путем количественной оценки гомологических отношений, основанных, например, на нейтральной теории Кимуры (синонимичной вариации области 3’-конца). Слева независимые линии инициируются в разное время при проникновении из космоса (из ●). Справа “Дерево” формируется на основании алгоритма, который предполагает прямые эволюционные взаимосвязи предшественников для всех гомологичных вариантов консервативного белка. Разветвленные группы F, G и HJJ представляют собой подлинные земные эволюционные диверсификации. A и B вымирают вскоре после проникновения из космоса. Адаптировано на основании концепций, изложенной у Hoyle and Wickramasinghe (1981), в частности рис. 6.3 и 6.4 на страницах 83-85. Основная концепция слева соответствует схеме прерывистого равновесия Элдриджа и Гулда (1972), Гулда и Элдриджа (1977). Обычная филогенетика любого типа, таким образом, работает на негласном предположении, что все гомологии связаны тем, что возникают только на Земле, игнорируя вклад космического эволюционного измерения.
< >Эволюционное происхождение ретровирусов позвоночныхИтак, это подводит нас к самой сути и важному уроку, который мы должны извлечь из этого анализа. В то время как все вирусы при внимательном рассмотрении оказываются
26 Ga/Giga-Annum- гигагод, единица измерения времени, равная одному миллиарду лет. Present - от англ. - Настоящее время (Прим.пер.)
чрезвычайно умными, ретровирусы (семейство Retroviridiae) входят в число наиболее сложных и комплексных из всех известных вирусов. Эти вирусы и их элементы (ферменты обратной транскриптазы, связанные с индуцированными мобильными ретро-элементами) теперь, по-видимому, являются важными вирусными двигателями основных эволюционных генетических изменений на Земле за последние несколько сотен миллионов лет (Wickramasinghe, 2012), и см. (Hoyle and Wickramasinghe,1979, 1981). Активность клеточной обратной транскриптазы (RT) также является основным молекулярным игроком в адаптивной соматической гипермутации генов антител (Franklin et al., 2004; Steele, 2016b; Steele and Lindley, 2017). Как только что обсуждалось, эндогенные ретровирусные векторы, а также активность RT, вероятно, вовлечена в ламарковскую передачу от сомы к зародышевой линии в иммунной системе (Steele, 1979; Steele et al., 1984; Steele et al., 1998; Steele and Lloyd, 2015; Steele, 2016a), включая формирование, дублирующие расширения и адаптивную перетасовку структур блочно-предковых гаплотипов генома человека и высших млекопитающих (Dawkins et al., 1999; McLure et al., 2013; Steele, 2014, 2015; Dawkins, 2015).
Основная генетическая и эволюционная особенность РНК-ретровируса заключается в том, что при продуктивной инфекции вирус обеспечивает создание двухцепочечной (ds) копии ДНК генома вирусной одноцепочечной (ss) РНК, и что она интегрируется в качестве провируса в геном клетки-хозяина. (Temin, 1974). В активном состоянии этот потенциально латентный провирус транскрибируется и производит вирусное потомство в инфицированной клетке для экспорта в другие клетки. Если клетка-хозяин является клеткой зародышевой линии, то интегрированный провирус может передаваться потомкам вертикально.
Ретровирусы могут инфицировать широкий спектр клеток животных, а полученные интегрированные провирусы могут оставлять свой след в виде “геномных окаменелостей”, называемых эндогенными ретровирусами (ERV). Эти последовательности ДНК, таким образом, являются пережитками прошлых и, вероятно, вирулентных ВИЧ-подобных эпидемий, в результате чего выжившие, частично невосприимчивые к непосредственному ВИЧ-подобному варианту, имеют геномную интеграцию в своих клетках зародышевой линии (что приводит, как обсуждалось, к дупликативному расширению и диверсификации функциональных блоков предковых гаплотипов (Dawkins et al., 1999; Dawkins, 2015).
Сейчас эти области ретровирусных исследований особенно популярны, благодаря большому количеству данных в общедоступных базах данных. В настоящее время обнаружено множество древних ERV27, что позволяет исследовать раннюю эволюционную историю ретровирусов с помощью биоинформатических методов. До недавнего времени предполагалось, что самые старые оценки возраста, полученные на основе анализа ретровирусов и ERV млекопитающих, датируются самым началом облучения млекопитающих и составляют примерно 100 миллионов лет. К настоящему времени, благодаря всеобъемлющему коэволюционному филогенетическому анализу (Aiewsakun and Katzourakis, 2017) стало ясно, что эндогенные ретровирусы костных рыб представляют собой модель совместного видообразования со своими хозяевами (но это еще не так ясно в отношении эндогенных ретровирусов амфибий). Однако общая коэволюция
27 Endogenous retroviruses - эндогенные ретровирусы. (Прим.пер.)
видообразования ERV с известными временными отрезками появления позвоночных видов их хозяев является очень убедительной. Возможные другие древние межвидовые и межклассовые вирусные трансмиссии происходили с низкой скоростью (включая лопастеперых рыб, акул и лягушек), однако анализ выявил две основные линии ERV у лучеперых рыб - одна линия из которых получила два новых дополнительных гена в своих гораздо более крупных геномах (~20 килобайт, или в два раза больше, чем у существующих ретровирусов млекопитающих). Таким образом, эта основная ретровирусная линия (и ретровирусы в целом), по-видимому, имеет древнее морское происхождение, которое “…возникло одновременно, если не раньше, с их челюстными позвоночными хозяевами >450 миллионов лет назад в Ордовикский период ранней Палеозойской эры” (Aiewsakun and Katzourakis, 2017). Это очень важный результат, и авторы приходят к выводу, что “… наш анализ дает как филогенетические, так и временные доказательства, что ретровирусы появились в океане одновременно со своими позвоночными хозяевами ~ 460–550 млн. лет назад, в раннюю Палеозойскую эру, если не раньше”.
Конечно, это относит даты появления этих сложных вирусных генных векторов - и ассоциируемое с ними воздействие эволюции, управляющие генами, - как раз ко времени около или до Кембрийского взрыва. Определенно они возникли еще до более позднего эволюционного появления гораздо более сложных видов животных и растений, ныне существующих на Земле. Другими словами, теперь мы можем привести убедительное научное доказательство того, что ключевая особенность генетических информационно-насыщенных систем, позволяющая создавать более сложные организмы, уже была здесь, на Земле, до фактического возникновения последующего более сложного земного многообразия. Это является противоречием, если мы примем во внимание нашу только что обсужденную точку зрения, согласно которой все вирусы, и особенно ретровирусы, являются “отражением в микрокосмосе генетической регуляторной структуры хозяина”, то есть, “тип сжатого модуля генетической информации, имеющего отношение к самой способности клетки расти и делиться, чтобы производить клетки-потомки, и развиваться” (Рис. 4).
- Кембрийский взрыв
Хорошо известно, что явление массового вымирания, или явления, произошли в конце Эдиакарского периода около 542 миллионов лет назад. Это период непосредственно предшествовал Кембрийскому взрыву, и, учитывая масштаб массового вымирания, можно предположить, что наша Солнечная система проходила через гигантское молекулярное облако, которое сместило несколько долгопериодических комет Облака Оорта во внутреннюю часть Солнечной системы, спровоцировав их столкновения с Землей (Hoyle and Wickramasinghe, 1981, 1993). Не нужно обладать богатым воображением, чтобы предположить, что предшествующие Кембрийскому взрыву события массового вымирания соотносились с воздействием несущей жизнь гигантской кометы (или комет) и последующим заселением Земли новыми клеточными организмами и вирусными генами космического происхождения (Hoyle and Wickramasinghe, 1979, 1981). Фактически, в начале Кембрийского периода действительно мог существовать обильный поток осколков
комет, послуживший причиной множественных столкновений с Землей на протяжении примерно 25 миллионов лет.
Явление столкновений с кометами характерно не только для докембрийского периода. Существуют и другие свидетельства, когда столкновения с космическими болидами совпадали с явлениями массового вымирания и диверсификации (например, на границе K-T 65 миллионов лет назад28, отмеченной вымиранием динозавров и увеличением количества мелких млекопитающих), а также хорошо установленная периодичность протяженностью в 26 миллионов лет (Галактический прилив - цикл Холмса), когда кометные удары совпадали с явлениями массового вымирания (Clube et al., 1996) и эволюционной диверсификацией (Napier et al., 2007). Экзотические аминокислоты в пограничных породах K-T и рост микрогрибков также указывают на появление нового генетического материала в результате столкновения (Wallis, 2007).
В этом отношении работа Мартина Бразьера, доказывающая периодичность в 20-30 миллионов лет в коэволюции дырочников29 и водорослей, может быть интерпретирована с Космической, а не с Земной точки зрения, которую предпочитал сам Бразьер (Brasier, 2012, стр.156 -157), когда указывал, что периодичность в 20-30 миллионов лет - это свойство систем крупных организмов быть уязвимыми к небольшим изменениям. Хотя теория и анализ Х.-В. предполагают довольно значительное изменение окружающей среды, вызванное воздействием Космоса, наш вывод согласуется с недавним взглядом Ноубла о том, что “Причинность множественна в биологии” (Noble, 2011). В этом контексте следует также подчеркнуть, что “прерывистое равновесие” Элдриджа-Гулда (рис.6) часто представляется как объяснение наблюдаемых фактов эволюции, но это не так. Это всего-лишь использование слов для описания поразительного феномена, который сам по себе не имеет убедительного объяснения в рамках биологии, ориентированной на Землю.
Само собой разумеется, что Тихоходки, микросегментированные крошечные эукариотические животные, появившиеся в Кембрийский период, представляют собой серьезный вызов традиционному неодарвинистскому мышлению (см. ниже суммарную справку о Тихоходках из Википедии и дополнительные подробные ссылки на сайте Википедии https://en.wikipedia.org/wiki/Tardigrade, а также данные, приведенные Jönsson et al., 2008). Перечень “существующих” свойств космической выносливости полностью соответствует “эволюционным естественным селективным явлениям”, которые действуют на тихоходок, развивающихся во внеземных космических условиях.
Эти свойства несовместимы с известными условиями чисто земного естественного отбора ни сейчас, ни 500 миллионов лет назад. Приемлемые доказательства существования космической панспермии могут полностью основываться лишь на одном этом примере. Однако, как уже довольно подробно обсуждалось, существует множество других подобных живых примеров (устойчивые к условиям космоса виды бактерий и их споры), и Тихоходки, будучи экстремальным примером, кажется, лишь подтверждают это правило (Hoyle and Wickramasinghe, 1981, 1993).
28 Граница K-T (от англ. Cretaceous–Tertiary) - граница Мелового и Третичного периодов. (Прим.пер.)
29 Дырочники - русское название Фораминифоров (лат. Foraminifera). (Прим.пер.)
Опубликованные недавно результаты эксперимента, проведенного доктором Томасом Лейя (Leya et al., 2017 в Институте клеточной терапии и иммунологии им. Фраунгофера (IZI)30 в Потсдаме, Германия, в сотрудничестве с немецкими и международными партнерами, соответствуют этой точке зрения.
Тихоходки (длиной 0,5мм во взрослом состоянии) образуют тип Tardigrada, часть супертипа Ecdysozoa, - древнюю группу, история окаменелостей которой уходит на 530 млн. назад в Кембрийский период. Описано около 1150 видов тихоходок. Их можно найти по всему миру, от Гималаев на высоте выше 6000 м (20 000 футов) до глубоководного океана на глубине ниже 4000 м (13 000 футов), и от полярных регионов до экватора. Эти водные восьминогие сегментированные микро-животные были найдены повсюду, от горных вершин до морских глубин, от влажных тропических лесов до Антарктиды. Это самое стойкое животное из всех известных, которое способно:
- выживать в экстремальных условиях, являющихся фатальными для почти всех других известных форм жизни;
- выдерживать высокие температуры от 1K (-458F%/-272°C), близкую к абсолютному нулю, до примерно 420K (300°F/150°C);
- давление примерно в шесть раз больше, чем давление в самых глубоких океанических желобах, ионизирующих радиацию в дозах, в сотни раз превышающих смертельную дозу для человека;
- вакуум космического пространства;
- обходиться без еды и воды более 30 лет, высыхая до состояния, когда они содержат всего около 3% (или меньше) воды, достаточное только для того, чтобы регидратировать, питаться и размножаться. (Источник: Википедия)
Группа привела доказательства того, что два вида водорослей выжили после пребывания в течение 16 месяцев на внешней стороне Международной космической станции (МКС), несмотря на резкие колебания температуры и космический вакуум, а также значительное ультрафиолетовое и космическое излучение
30 нем: Fraunhofer-Institut für Zelltherapie und Immunologie (IZI) (Прим.пер.)
(http://www.algaeindustrymagazine.com/algae-survive-outside-space-station/). Логические последствия этих данных для космической биологии очевидны - фактически мы утверждаем, что такие открытия имеют гораздо больший смысл в рамках парадигмы космической эволюции Х.-В. о космической биологии - панспермии (Приложение A на стр 40).
- Последствия кембрийского взрыва и данные о ретровирусах
До того, как стал доступен способ обширного секвенирования ДНК, было разумно предполагать, что случайные ошибки копирования в последовательности гена могут со временем привести к появлению новых черт, строения тела и новой физиологии, которые могли бы объяснить всю эволюцию. Однако данные, которые мы здесь рассмотрели, ставят под сомнение эту точку зрения. Предполагается, что Кембрийский взрыв многоклеточной жизни, произошедший 0,54 миллиарда лет назад, привел к внезапному появлению практически всех генов, которые впоследствии были преобразованы в чрезвычайно широкий спектр многоклеточных форм жизни в виде тихоходок, кальмаров, осьминогов, дрозофил, людей и многих других.
Экспрессия целого нового набора ретровирусных генов (предполагаемого внешнего происхождения), по-видимому, происходила в основном за счет геномных перестроек, последовавших за интеграцией ERV. Это была именно логика эволюции, которую предложили покойный Сэр Фред Хойл и один из соавторов этой статьи (N.C.W.) еще в 1981 году. Они утверждали, что ошибки копирования существующих генов не могут в среднем производить новые гены с функциональной полезностью. Они также указали, что по аналогии с компьютерным программированием ошибки, возникающие при копировании компьютерного кода, не приведут к расширенным или новым возможностям, а в подавляющем большинстве случаев приведут к деградации исходной программы. Поэтому доказывалось, что, логически новые гены для эволюции должны поступать посредством проникновения внеземных вирионов и других микроорганизмов.
- Космическая биология и зарождение человечества
Так, Хойл и Викрамасингхе (Hoyle and Wickramasinghe,1979, 1981, 1986) выстраивали свои предположения и прогнозы, используя имеющуюся в то время информацию (Рис.1 и Рис.2) о массовом исчезновении видов, об астрономических инфракрасных спектрах межзвездных и кометных тел, и позднее - на основе известных зафиксированных свидетельств об ударных кратерах Земли31 и о регулярных кометных ударах (Clube et al., 1996). Эти данные помогли им сделать вывод, что микроорганизмы и популяции вирусов в кометах и связанных с ними космических болидах, по-видимому, регулярно доставляли живые системы (организмы, вирусы и семена) на Землю с момента ее образования и продолжают это делать. Безусловно, Земля должна была быть “гостеприимным” местом и, следовательно, восприимчивой к этим живым системам, принесенным кометами. Это
31 См. Список ударных кратеров Земли — Википедия (wikipedia.org). (Прим.пер.)
наводит на одно из вероятных объяснений того, почему взрывы, подобные кембрийскому, не происходили на протяжении 3500 миллионов лет, предшествующих периоду, начавшемуся 500 млн лет назад, несмотря на вероятное прохождение нашей Солнечной системы через множество других галактических молекулярных облаков в этот временной интервал при том, что галактический год нашего Солнца на его орбите вокруг Галактического центра составляет примерно 240 миллионов лет (Hoyle and Wickramasinghe, 1993; JT Wickramasinghe et al., 2010). Иначе, это означает, что огромный поток космических отходов, состоящий из сложных эукариотических организмов (растений, животных, грибов), прибывал на Землю на протяжении 25 миллионов лет в самом начале Кембрийского периода, редкого и значительного явления космического засеивания, которое произошло около 500-550 млн. лет назад, а не в предшествующие им 2000–3000 млн. лет. Поэтому в силу данной необходимости наша дискуссия здесь ведется именно в масштабах космического пространства-времени.
Таким образом, за последнее десятилетие увеличилось не только количество доказательств, согласующихся с гипотезой Х.-В. о космической биологии, но и значительно окрепла их обоснованность (Napier et al., 2007; Rauf and Wickramasinghe, 2010; Wickramasinghe, 2010, 2011, 2012, 2015a, 2015b; Smith, 2013; Wickramasinghe, Smith, 2014). Мы убеждены в том, что необъятная космическая биология является наиболее вероятной движущей силой эволюционного многообразия жизни на Земле, и недавнее исследование П.Эюсакана и А.Кацуракиса (Aiewsakun and Katzourakis, 2017) является важнейшим звеном доказательства, которое идеально вписывается и соответствует мозаике этой картины.
Поэтому дарвиновская эволюция и ее различные недарвиновские наземные движущие силы (Wickramasinghe and Steele, 2016), скорее всего, явились результатом непрерывного поступления новых вирионов и микроорганизмов из космоса, а те в свою очередь стали причиной генетического воздействия, записанного по всему нашему геному (Wickramasinghe, 2012). В самом деле, можно привести веские доводы в пользу эволюции гоминидов, включая продолжительные циклы вирусных пандемий, каждая из которых подводила эволюционную линию к самому краю пропасти полного вымирания. Наиболее важные гены, имеющие отношение к эволюции гоминидов, как и всех видов растений и животных, во многих случаях, по-видимому, имеют внешнее происхождение и переносятся по галактике преимущественно в виде информационно-насыщенных вирионов. В некоторых случаях можно представить себе - причудливые, как это может показаться, - многоклеточные формы жизни, которые были образованы на ледяной комете или планетарном теле, чтобы огромные ледяные болиды транспортировали их в виде замороженных яйцеклеток, эмбрионов или семян (Tepfer and Leach, 2017), а затем мягко доставили на Землю (Frank and Sigwarth, 2001; Snyder, 2015); безусловно, земные свидетельства наличия на нашей планете широко распространенной ледяной биосферы вполне убедительны и соответствуют такой картине (Priscu and Christner, 2004; Fox, 2014; Christner et al., 2014). Вполне вероятно, что в нагретых и жидких подповерхностных недрах комет или планетных лун, таких как Европа Юпитера и Энцелад Сатурна (Hoover, 2011; Snyder, 2015), неизбежно должны будут существовать клеточные резервуары для репликации вирусов.
- Причины эпидемий и пандемий: новый принцип вирусологии
То, что человечество поразило множество болезней, эпидемий и пандемий, не подлежит сомнению. ВИЧ/СПИД - один из самых распространенных и недавних. Ретровирусное заболевание, передающееся половым путем, и, очевидно, поражающее иммунную систему и вызывающее ее последующее разрушение, возникло совершенно неожиданно, - и это не было научной фантастикой! Крупные центры мониторинга, такие как CDC32 в Атланте и Пирбрайт33 в Великобритании, были застигнуты врасплох и оказались абсолютно неподготовленными. Очевидно, что и пандемия Испанского гриппа 1918 года была столь же разрушительной и столь же неожиданной. В высшей степени необычная эпидемиология пандемии вируса испанского гриппа 1918 года - в эпоху, предшествующую авиапутешествиям, - была проанализирована очень подробно, как и эпидемиологические мельчайшие детали недавних эпидемий гриппа (Hoyle and Wickramasinghe, 1979). Эти исследования полностью опровергают популярную и традиционную биомедицинскую точку зрения, согласно которой инфекционные заболевания передаются главным образом от человека к человеку. Это чрезвычайно убедительное свидетельство было подробно проанализировано и задокументировано в книге Хойла и Викрамасингхе (Hoyle and Wickramasinghe, 1979). Многие коллеги N.C.W., соавтора этой обзорной статьи, утверждают, что эти неоспоримые свидетельства должны быть проанализированы абсолютно всеми биологами и учеными-биомедиками, желающими понять наиболее вероятные движущие силы многих очевидно неожиданных (и полностью необъяснимых) инфекционных эпидемий, которые случаются регулярно: многие с годовой периодичностью или периодичностью, совпадающей с 11-летними циклами солнечной активности (Wickramasinghe et al., 2017). В случае гриппа внезапное появление множественных, но очаговых зон заражений (многие из которых, как указывалось, возникли до появления авиаперелетов) не может быть объяснено простыми моделями передачи инфекционного заболевания от человека к человеку. Тем не менее, их возникновение можно полностью объяснить причинами многократных космических ударов или попадания из космоса в совершенно разные точки мира, и это зависит от таких превратностей, как погода, топография, география, и, в частности, существенно коррелирует с периодическими вспышками 11-летнего Солнечного цикла (Hope-Simpson, 1978; Hoyle and Wickramasinghe, 1979; обзор в Qu, 2016; Wickramasinghe et al., 2017). Что касается последней корреляции, “... ожидается, что пики солнечной активности будут способствовать спуску заряженных молекулярных агрегатов (включая вирусы) из стратосферы на уровень земли ... вдоль силовых линий магнитного поля, соединяющих Солнце и Землю” (Wickramasinghe, 2015a, стр. 81; Wickramasinghe et al., 2017).
Можно утверждать, что и доказательства, изложенные в теории Х.-В., и личные контакты при общении между людьми являются причинами возникновения эпидемий, причем некоторые эпидемии больше соотносятся с теорией Х.-В., а некоторые - с межличностными контактами. Очевидное подтверждение этого - все мы имели опыт простуды, заразившись от кого-то другого, и можем точно вспомнить, когда и с кем из
32 CDC - Centers for Disease Control and Prevention (USA) - Центры по контролю и профилактике заболеваний в США. (Прим.пер.)
33 Pirbright Institute - Институт Пирбрайта, исследовательский институт в Англии, являющийся частью Совета по исследованиям биотехнологий и биологических наук при правительстве Великобритании. (Прим.пер.)
переносчиков этого заболевания мы пересекались. Этот опыт, конечно же, дает всем нам предвзятое мнение о том, что “единственный способ заразиться вирусом - это прямо или косвенно от кого-то другого”. Однако именно это широко распространенное мнение прямо опровергается подробным анализом причин внезапно возникающих эпидемий гриппа Хойла и Викрамасингхе (Hoyle and Wickramasinghe, 1979. В частности, см. главу 6 “Анатомия эпидемии” и главу 5, содержащую критический анализ пандемии гриппа 1968 года), когда подобные предполагаемые близкие физические контакты лицом-к-лицу не могут объяснить схемы распространения гриппа (т. е. распространение вируса через телесные жидкости, такие как слюна, кровь, сперма, или распыленные вблизи вашего лица взрывоопасные аэрозоли). В других случаях, таких как Vibrio cholerae34, бактерия передается через воду. Зараженная вода, попадая прямо в рот и пищеварительный тракт восприимчивых жертв, может явиться непосредственным физическим переносчиком вируса и вызвать Холерную диарею. Вспышки пищевого отравления Сальмонеллой - еще один известный пример этого типа. Таким образом, хотя очевидно, что существует сочетание видов передачи вирусов по теории Х.-В. и от человека к человеку, реальная причина действительно зависит от изучаемого заболевания. Но если вирусное заболевание возникает неожиданно, как взрывоопасная волна, и имеет масштаб крупной эпидемии или пандемии, то “воздействие из космоса” должно быть одной из возможных стандартных первопричин, подпадающих под подозрение. Все это подробно обсуждается в книге Хойла и Викрамасингхе (Hoyle and Wickramasinghe, 1979). Несмотря на многолетние исследования вакцин против гриппа и более глубокие знания о местах скоплений вируса птичьего гриппа, факт того, что сезонный грипп и эпидемии/пандемии плохо изучены, по-прежнему остается актуальным. Это невежество касается как появления новых антигенно непредсказуемых штаммов, так и фактического пути передачи болезни во время вспышек и эпидемий (Paules et al., 2017).
Поскольку сейчас мы рассматриваем “космический генофонд” с точки зрения все пронизывающей космической биологии, это означает, что многие новые возникающие болезнетворные микроорганизмы и вирусы будут вступать в филогенетические отношения с существующими вирусными видами и родами (Рис. 6). В качестве первого быстрого объяснения мы можем предположить, что вирусы пришли из космоса, - даже если есть и другие правдоподобные земные объяснения первопричины их появления, - то есть космическая причина должна быть одной из совокупности причинно-следственных объяснений, и должна быть либо учтена, либо исключена.
Но что делать с космическим вариантом? На данном этапе нашим ответом может быть только долгосрочная стратегия наблюдения, нацеленная на накопление знаний о предупреждающих признаках надвигающихся ударов патогенных кометных обломков. Это означает организованное размещение мониторинговых космических аппаратов и регистрирующих станций наряду с более обширными и систематическими методами отбора проб при помощи аэростатов (Wainwright et al., 2014, 2015a, 2015b), а также внедрение сложной системы наблюдения, прототип которой, известный как “Щит Хойла”, находится на самой ранней стадии разработки (Smith, 2013).
34 Холерный вибрион - вид грамотрицательных факультативно анаэробных подвижных бактерий рода вибрионов. (Прим. пер.)
- Смена парадигмы
По нашему твердому убеждению, Человечество сейчас приблизилось к моменту смены исторической парадигмы и нашего понимания как эволюции жизни на Земле, так и истоков многочисленных пандемий, которые еще совсем недавно привели человечество к огромным потерям. По сути, мы все внесли свой непосредственный вклад в качестве соавторов этой статьи, потому что, с нашей взвешенной точки зрения при сопоставлении широкого спектра научных данных, все научные и общественные доказательства, по-видимому, указывают в одном направлении: в сторону повсеместно присутствующей космической биологии, опосредованной преимущественно кометными трансферами, которая и является движущей силой жизни на Земле. Мы считаем, что признаки этих изменений в настоящий момент настолько очевидны, что один из самых больших разворотов в истории науки готов случиться прямо сейчас (Приложение C на стр. 46).
Если вы думаете, что наша позиция здесь надуманная или даже паникерская, мы процитируем слова великого профессора Корнельского университета, покойного Томаса Голда. Голд, дальновидный и изобретательный астроном и геофизик (Gold, 1992, 1999), который совершил далеко не один научный разворот на пути своей долгой карьеры, наиболее заметный из которых был связан с науками о Земле в середине-конце 1960-х годов и резкой сменой курса в сторону теории дрейфа континентов Альфреда Вегнера 1912 года. В 1989 году Том Голд писал:
“Как на самом деле выглядит процедура рецензирования? Как это происходит на самом деле? Если, например, в начале 60-х или конце 50-х годов подавалось заявление, в котором излагалось, что человек хотел бы исследовать возможность небольшого перемещения континентов, оно было бы отклонено абсолютно мгновенно и без вопросов. Это было чепухой сумасшедшего, и уже давно считалось ненужным. Вегенер, конечно, был абсолютно чокнутым, и все это знали, и у вас не было бы никаких шансов. Шесть лет спустя вы не могли опубликовать статью, которая ставила бы под сомнения дрейф континентов. Стадо качнулось - но по-прежнему осталось устойчивым и высокомерным стадом”. Проверьте это на http://amasci.com/freenrg/newidea1.html.
Похожий пример был связан с широко распространенным официальным отрицанием реальности существования метеоритов во времена Лавуазье (1768 г.), который “отвергал рассказы очевидцев о падении метеоритов по причине здравого смысла, согласно которому “камни не могут падать с неба, поскольку на небе камней нет”“ (Haynes, 1980).
Тезис Хойла-Викрамасингхе о влиянии повсеместно присутствующей космической биологии, сформировавшейся на Земле в основном при помощи комет, полностью соответствует экспериментальным и обсервационным данным, накопленным за последние 40-50 лет с момента первого упоминания этой идеи. Хотя полное принятие этих многофакторных и специфических идей произойдет со значительным опозданием, они все равно станут великим научным достижением, и мы надеемся и верим, что новые сторонники нашей теории проявят доброту, внимательность и уважение по отношению к тем, кто являлись пионерами в этой области, и признают первостепенное значение их научной деятельности.
Таким образом, учитывая всю лавину данных из всевозможных областей, каждая из которых указывает на повсеместное присутствие космической биологии, подразумевающей происхождение жизни вне Земли, перманентное нежелание научного сообщества признать этот факт может показаться странным. Тем не менее, как ясно указывает Том Голд - и все мы знаем о силе воздействия такого подхода в нашей повседневной жизни - “групповое мышление” и безопасность “бега со стадом” являются мощными движущими мотивационными силами как в науке, так и в обществе (Gold, 1989). Эти силы довольно иррациональны (с научной точки зрения), но очень мощны в социальном и культурном плане. Однако давно назревший сдвиг научной парадигмы от биологии, ориентированной на Землю, к космической жизни будет иметь серьезные последствия, выходящие далеко за рамки науки (Wickramasinghe and Tokoro, 2014a, 2014b). Несомненно, только в рамках биомедицинских наук прямое следствие филогенетического анализа, основанного на данных ДНК или аминокислотных последовательностей, потребует фундаментальной переоценки (сравните левую и правую эволюционные концептуальные траектории на рис.6). Однако последующие за этим потрясения в сфере политики, религии и человеческого самоосознания, вероятно, будут еще более драматичными и глубокими, чем те, которые произошли за последние 500 лет.
Подводя итоги, следует признать, что в конечном итоге вся эволюция контролировалась и продолжает контролироваться космическими организмами, микробами и вирусами. Важно не допустить, чтобы Наука была задушена господством догматического авторитета, стремящегося ограничить ее прогресс узкими консервативными направлениями. Нынешняя ситуация поразительно напоминает средневековье в Европе и эпициклы Птолемея, которые задержали принятие планетной системы с центральным положением Солнца более чем на столетие (Приложение C на стр. 46).
Имеющиеся данные свидетельствуют о том, что мы пришли из космоса, мы сделаны из вирусных генов, и в конечном итоге наше эволюционное наследие в полной мере вернется в космос. Это завершит вторую и последнюю фазу коперниканской революции, которая началась более полувека назад.
Конфликты интересов
Авторы заявляют об отсутствии конфликта интересов.
Вклад авторов
Разработка концепции: Эдвард Дж.Стил, Н.Чандра Викрамасингхе, Джон П.Коглан, Бриг Клайс, Дженсуке Токоро, Джон А.Шустер.
Сбор данных и публикаций с 1970-х по 2017 год: Эдвард Дж.Стил, Ширван Аль-Муфти, С.Дж.Кулсон, Реджинальд М.Горджински, Бриг Клайс, Годфрей Луис, Кит Р.Оливер, Цзянвэнь Цюй, У.Е.Смит, Дуэйн П.Снайдер, Брент Дж.Стюарт, Дженсуке Токоро,
Кристофер А.Таут, Милтон Уэйнрайт, Дэрил Х.Уоллис, Джейми Уоллис, Макс К.Уоллис, Д.Т.Викрамасингхе, Дж.Т.Викрамасингхе, Н.Чандра Викрамасингхе, Юншэн Лю.
Формальный анализ и интерпретация различных аспектов данных: Эдвард Дж.Стил, Ширван Аль-Муфти, С.Дж.Кулсон, Реджинальд М.Горджински, Бриг Клайс, Годфрей Луис, Кит Р.Оливер, Цзянвэнь Цюй, У.Е.Смит, Дуэйн П.Снайдер, Дженсуке Токоро, Кристофер А.Таут, Милтон Уэйнрайт, Дэрил Х.Уоллис, Джейми Уоллис, Макс К.Уоллис, Д.Т.Викрамасингхе, Дж.Т.Викрамасингхе, Н.Чандра Викрамасингхе, Юншэн Лю.
Текст - первоначальный вариант: Эдвард Дж.Стил, Дженсуке Токоро, Н.Чандра Викрамасингхе.
Текст - проверка и редактирование: Эдвард Дж.Стил, Ширван Аль-Муфти, Кеннет А.Августин, Роана Чандраджит, Джон П.Коглан, С.Дж.Кулсон, Судипто Гош, Марк Гилман, Реджинальд М.Горджински, Бриг Клайс, Годфрей Луис, Китсири Маханама, Кит Р.Оливер, Хулио Падрон, Цзянвэнь Цюй, Джон А.Шустер, У.Е.Смит, Дуэйн П.Снайдер, Джулиан А.Стил, Брент Дж.Стюарт, Роберт Тэмпл, Дженсуке Токоро, Кристофер А.Таут, Александер Унзикер, Милтон Уэйнрайт, Джейми Уоллис, Дэрил Х.Уоллис, Макс К.Уоллис, Джон Уэтералл, Д.Т.Викрамасингхе, Дж.Т.Викрамасингхе, Н.Чандра Викрамасингхе, Юншэн Лю.
Примечание к доказательству
Важнейшее предсказание космической теории о возникновении жизни Хойла-Викрамасингхе состоит в том, что новый генетический материал (ДНК/РНК) в форме бактерий и вирусов прибывает на Землю из космоса как постоянно, так и спорадически. Предполагается, что такой процесс расширяет развитие биологической эволюции, включая горизонтальный перенос генов сквозь галактические или космические измерения. Поиск таких поступающих бактерий и вирусов с использованием специально-оборудованного аэростата запущенного в стратосферу велся в течение почти двух десятилетий (Harris et al, 2002; Wainwright et al, 2003; Shivaji et al, 2009). Пробы стратосферного воздуха, собранные на высоте от 28 до 41 км, действительно выявили наличие микроорганизмов, но многие считают, что они, скорее всего, были подняты с поверхности Земли. Однако за последние месяцы ситуация кардинально изменилась. Т.В. Гребенникова и ее коллеги (T.V. Grebennikova et. al., 2018) подтвердили открытие нескольких видов микробов, соответствующих космической пыли, на внешних окнах Международной космической станции (МКС), причем загрязнение в источнике и в лаборатории исключено. Результаты ПЦР-амплификации с последующим секвенированием ДНК и филогенетическим анализом установили присутствие бактерий рода Mycobacteria и экстремофилов рода Delftia, среди прочих, связанных с отложениями космической пыли, находящихся на высоте около 400 км над земной поверхностью. Земное происхождение представляется маловероятным. Исследования Викрамасингхе и Рикрофт (Wickramasinghe and Rycroft, 2018) показали, что все возможные механизмы подъема этих организмов против силы тяжести на высоту 400 км в ионосферу на много порядков неэффективны.
Приложение A. Происхождение жизни и научные альтернативы космической панспермии Х.-В.?
“Давайте постараемся постичь то, что постижимо, и оставим то, что непостижимо, для благоговения в одиночестве ... “ Иоганн Вольфганг фон Гете
На этом этапе научного понимания нам необходимо отложить в сторону вопрос о фактическом биохимическом происхождении жизни: вопросы “где, когда и как” могут оказаться слишком сложными для ответа на основе имеющихся данных. Настоящая статья сосредоточена на доказательствах всепроникающей космической биологии и ее влиянии на возникновение жизни на Земле и ее дальнейшую эволюцию. Определенно, все попытки абиогенеза в лаборатории на Земле были безуспешными. На много порядков более вероятно, что этот процесс возник в одном из триллионов кометоподобных инкубаторов или водоносных планет (космические версии “теплых маленьких прудов” Дарвина) в очень ранний период роста Вселенной, возможно, 12 миллиардов лет назад (Wickramasinghe, 2015a), после чего продолжил вирулентное распространение посредством косвенного влияния на другие благоприятные для жизни места (планеты, луны, кометы) по всей Галактике, а затем взаимосвязанным и интерактивным образом по всему космосу, так как Вселенная расширяется. Сферическое Облако Оорта, источник возникновения долгопериодических комет во всей нашей Солнечной системе, помогает понять причинно-следственную связь и объяснить это предположение. Возможно, большинство из этих миллиардов комет возникли не из плоскости аккреции нашей Солнечной системы. Скорее всего, они появились в результате захвата комет силой притяжения нашего Солнца из других перемещающихся мимо солнечных систем (Levison et al., 2010).
По мнению одного из авторов этой статьи (B.K.)35, “многогранная и утонченная жизнь не может возникнуть (из небиологической) материи ни при каком сценарии ни в каком пространстве и времени, какими бы огромными они ни были. Если бы это было так, тогда вслед за типичным Большим взрывом потребовалось бы либо сверхъестественное вмешательство, либо вмешательство интеллекта (см. также Walker and Wickramasinghe, 2015). Строго научный способ решения этой дилеммы состоял бы в том, чтобы внести поправки или подкорректировать теорию большого взрыва с учетом факта присутствия жизни в вечном прошлом. В конце концов, теория большого взрыва относительно нова, и в нее все еще время от времени вносятся уточнения. Поэтому представляется неразумным навсегда отвергнуть целостный принцип и убедительное доказательство того, что жизнь произошла от жизни”.
Другой из наших соавторов (E.J.S.)36 смотрит на этот же сценарий несколько иначе: исходя из положений, опубликованных Хойлом и Викрамасингхе (Hoyle and Wickramasinghe, 1978b, 1979, 1981) о том, “является ли Вселенная бесконечным стационарным состоянием (непрерывная серия больших взрывов и сжатий) - как это было высказано Фредом Хойлом, Томом Голдом и Германом Бонди в 1948 году - или это просто обычный “Большой взрыв”? Последнее - это то, чего в настоящее время придерживается большинство
35 Бриг Клайс (Прим.пер.)
36 Эдвард Дж.Стил (Прим.пер.)
физиков-математиков и астрономов, но этот вариант ни в коем случае нельзя считать незыблемым (Tegmark, 2014). Обе альтернативы приводят в замешательство рациональный научный ум. Но если нам суждено сделать выбор, то мы должны учесть охватывающую весь космос взаимозависимую биологию, стационарное состояния Вселенной (или квазистационарное состояния Вселенной, Hoyle et al., 2000) и бесконечность времени, - все факторы, которые обеспечивают временную протяженность, необходимую для того, чтобы в триллионах маленьких теплых прудов по всему Космосу методом проб и ошибок и путем естественного отбора запустился процесс абиогенеза (чтобы объединить феноменально невероятную живую клетку воедино с тысячами интерактивных генов, управляющих процессами клеточной жизни в космическом масштабе). Таким образом, Хойл и Викрамасингхе (Hoyle and Wickramasinghe, 1981) пришли к выводу, что вероятность случайной сборки живых белков и нуклеиновых кислот настолько мала (1:1040000), что, наверное, потребовались бы “Бесконечная Вселенная” или сверхразумный Бог, чтобы произвести живое чудо, которое затем распространилось и эволюционировало бы в масштабе всего Космоса”. Размышления над этими глубокими обсервационными проблемами могут вполне объяснить, почему выдающийся философ-гуманист, покойный Энтони Флю (Antony Flew, 2007) честно и открыто поменял свои философские воззрения на прямо противоположные.
Однако, что касается непосредственного происхождения жизни на Земле, мы, учитывая все имеющиеся данные, полагаем, что не существует ни одной серьезной научной теории, которая могла бы составить альтернативу космической (кометарной) гипотезе Хойла-Викрамасингхе о панспермии как главной движущей силе жизни на Земле. Когда примерно в 1960 году Тома Голда публично спросили о имеющихся у него доказательствах, он, поразмыслив и, очевидно, полушутя, заявил: “Давным-давно на Землю приземлился космический корабль и разбросал живые клетки, которые с тех пор эволюционировали и упорно продолжают свое существование.” (Hoyle and Wickramasinghe, 1981, стр.30). В 1973 году этому придали значание Фрэнсис Крик и Лесли Орджел и опубликовали свою собственную гипотезу о “направленной панспермии”, основная идея которой состояла в том, что жизнь на Земле была засеяна три-четыре миллиарда лет назад запустившей свои космические корабли сверхразумной цивилизацией. Научным обоснованием для продвижения такой теории стало то, что необходимо было найти объяснение необычным фактам о чрезвычайной сложной, но при этом “универсальной” структуре генетического кода, - который действительно позволяет нам клонировать и экспрессировать человеческие гены в бактериях и наоборот - и лежит в основе всей современной биотехнологии, биомедицины, терапевтических и биомедицинских исследований. Объяснение Х.-В., на наш взгляд, является гораздо более обоснованным и научно-подкрепленным (скрупулезная модель).
Один из критически-настроенных соавторов нашей статьи (K.A.A.)37 высказал предположение, что и теория Х.-В., и жизнь, возникающая из неживого на нашей планете, возможно, верны. Другими словами, по мнению K.A.A.: “Жизнь могла возникнуть из неживого разными способами и в разных местах, в том числе и здесь, на Земле”. Таким образом, панспермия полностью согласуется с идеей возникновения живых организмов. В недавней книге “Вопрос жизни” Ник Лейн (Nick Lane, 2015) приводит возможную теорию
37 Кеннет А.Августин (Прим.пер)
происхождения живого из неживого на Земле, и предполагает, что жизнь могла возникнуть аналогичным образом на независимой основе во многих местах Вселенной. Хойл и Викрамасингхе уже подробно проанализировали эту возможность земного происхождения. C их точки зрения, этот сложный вариант версии о самопроизвольном зарождении, которая была опровергнута Луи Пастером в XIX веке. Более того, такой взгляд лишь бесконечно усугубляет изначальные несоответствия, поскольку предусматривает зарождение жизни только в одном месте. Он также не принимает во внимание тот общепризнанный современный факт, что только в одной галактике существуют сотни миллиардов пригодных для жизни планет со средним расстоянием между ними порядка нескольких световых лет. Поэтому обмен биотическим материалом между соседними планетами и, следовательно, единой связанной биосферой, представляется неизбежным. Учитывая все имеющиеся доказательства, мы предлагаем рассматривать внеземное зарождение жизни на ранних этапах формирования Космоса как наиболее вероятную версию (опять-таки, модель Х.-В. является более скрупулезной).
Наряду с этим, очень странным кажется то, что Ник Лейн в своей книге “Вопрос жизни” (Nick Lane, 2015) отвергает идею панспермии как не относящуюся к делу: “Панспермия совершенно не соответствует этим принципам (этапам, которые он описывает как важные в процессе происхождении жизни посредством абиогенеза) и поэтому не имеет отношения к делу…” (стр.94). Такой взгляд полностью упускает из виду смысл существующей всепроникающей внеземной биологии в нашей Солнечной системе и, почти наверняка, - согласно данным о межзвездном инфракрасном поглощении - во всем Космосе. Об этом Ник Лейн, как кажется, просто забыл. Поразительно противоречиво, но в этой же книге (стр. 55-56) Лейн рассказывает о тихоходках (инфосправка из Википедии) и подробно описывает их космические характеристики! Поэтому ключевой вопрос - это кратко изложенные в этой статье многочисленные свидетельства о повсеместном присутствии космической биологии, c которыми человечеству теперь придется иметь дело. Эти знания позволят нам лучше понять текущий эволюционный процесс, а также причины возникновения новых спонтанных эпидемий и болезней у наших домашних животных и растений, а также у людей.
Некоторые авторы научно-популярных книг, такие как Ник Лейн из Университетского колледжа Лондона, сочли модным отвергнуть гипотезу Хойла-Викрамасингхе буквально одной строкой. Это разочаровывает, потому что в его книге содержится много нетрадиционных и новаторских размышлений о вероятных сценариях возникновения жизни не только на Земле, но и во всем Космосе. Нет никакой необходимости пренебрегать очевидными данными, которые нас окружают, лишь по причине космической биологии. Хотя причинно-следственная связь абиогенеза для Земли представляется маловероятной, его наглядные и хорошо обоснованные сценарии для пористых щелочных гидротермальных (на средних широтах) источников срединно-океанических хребтов как мест раннего вероятного биоэнергенеза (ранние митохондриально-подобные протонно-градиентные мембранные энергетические системы) могут быть протестированы в лаборатории. Более того, множество экспериментов, которые дают почву для размышлений Нику Лейну (а также его сотрудникам, таким как Билл Мартинс, и его аспирантами), необходимо поощрять и финансировать. Рано или поздно, после продолжительных попыток Поппера и устранения ошибок в его многочисленных шагах на пути к самовоспроизводящимся энергетическим живым системам, мы не сомневаемся, что
человечество во главе с такими людьми, как Ник Лейн, в конечном итоге проведет успешный эксперимент по абиогенезу. Он будет основан на комплексном понимании прошлых неудач и продемонстрирует “происхождения жизни” из простых космических исходных материалов (H2, CO2, CO, силикатов, фосфатов, железо-серных агрегатов, H2O и т.д.) в пробирке. Для человечества, это действительно был бы эксперимент, изменивший эпоху. Но на данный момент у нас есть огромное количество скупых данных о внеземной жизни вокруг нас, и мы должны учесть их и проанализировать все последствия.
Другой соавтор и физик-теоретик (A.Z.)38 развивает сценарий Тома Голда и рассматривает вероятность космического происхождения следующим образом: “Поскольку идея космического происхождения жизни еще не принята в области эволюционной биологии, она пока что не завоевала популярность среди ученых общего профиля, однако это может измениться. В своей книге 2014 года “Искусственный интеллект” оксфордский философ Ник Бостром (Nick Bostrom, 2014) утверждает, что человечество рано или поздно разовьет формы интеллекта, значительно превосходящие человеческий уровень. Произойдет ли это в краткосрочной перспективе (с помощью искусственного интеллекта, генерируемого новыми алгоритмами в существующем оборудовании) или в более отдаленном будущем, когда нанотехнологии позволят создавать копии человеческого мозга, не имеет значения для нашего обсуждения здесь - в астрономических масштабах времени это произойдет в мгновение ока в обоих случаях. Такой суперинтеллект легко сможет (и, вероятно, захочет) построить космический корабль (или более мелкие единицы), который сможет эффективно распространять примитивные формы жизни в других регионах Млечного Пути вдали от Солнечной системы, то есть путем панспермии. Временными рамками, необходимыми для этого, будут тысячи, а не миллионы лет, то есть ничтожно малый срок. Так что это, безусловно, приемлемая гипотеза о том, что на такой сверхразумной планете могло бы быть от 10 до 100 нисходящих цивилизаций. Однако это означало бы, что у нас есть априорная вероятность того, что мы имеем космическое происхождение на 90-99 процентов - это то, на что указывает простой байесовский анализ39 после всего лишь одного “поколения””.
A.Z. продолжает: “Примечательно, что тезис Бострома до сих пор не подвергся сомнению
- похоже, что никаких серьезных контраргументов не существует (например, см. Википедию, “Суперинтеллект”). Тем не менее, это имеет глубокие последствия для нашего обсуждения здесь. У науки иногда бывает шизофрения: гипотеза может быть интересной и убедительной в одной области (здесь: нейробиология), тогда как в другой области считается, что ее не стоит обсуждать (эволюционная биология)”.
Но очевидно, что есть один очень важный процесс, обнаруженный и разработанный Иреной Косич и ее коллегами: распознавание молекулярного резонанса между рецепторами макромолекул и их лигандом (белки, нуклеиновые кислоты), которое необходимо интегрировать во все размышления о том, как, прежде всего, в мире могла возникнуть и продолжает развиваться жизнь. Это опосредованное электромагнитным
38 Имя по инициалам в списке авторов статьи не найдено. Возможно, имеется в виду Александр Унзикер. (Прим.пер.)
39 Байесовский анализ — статистический вывод, в котором свидетельство и/или наблюдение используются, чтобы обновить или вновь вывести вероятность того, что гипотеза может быть верной; название байесовский происходит от частого использования в процессе вывода теоремы Байеса. (Прим.пер.)
излучением межмолекулярное распознавание и функциональная активация до фактического связывания, что важно для всех живых систем и должно учитываться во всех моделях и экспериментах по абиогенезу (Cosic, 1994); Cosic et al., 2015, 2016; Cosic and Cosic, 2016).
С быстро растущим числом экзопланет, обнаруженных в обитаемых зонах долгоживущих красных карликов (Gillon et al., 2016), нельзя игнорировать перспективы генетического обмена между жизнеспособными планетами земного типа. На наш взгляд, идея возникновения жизни de novo40 в нескольких местах в процессе абиогенеза in situ41 кажется надуманной и лишь усугубляет трудность, связанную с единым происхождением на Земле. Внутренние данные о земной биологии, которые мы обсуждали в этой статье, предполагают, что весь ансамбль обитаемых планет в галактике составляет единую взаимосвязанную биосферу. С этой точки зрения ожидается, что формы жизни в других местах будут демонстрировать сливающуюся структуру генотипов и фенотипов, подверженных естественному отбору (дарвиновскому и ламарковскому) в пределах индивидуальных планетарных сред обитания. Таким образом, недавнее объявление о семи “обитаемых” планетах вокруг звезды Trappist-1, удаленной от нас на 40 световых лет, является весьма своевременным (Gillon et al., 2016) и имеет отношение к тезису, обсуждаемому в настоящей статье. Планируются дальнейшие исследования для поиска спектральных сигнатур молекулярного кислорода и метана в планетных атмосферах как индикаторах жизни; хотя такие молекулы (и многое другое) действительно были обнаружены на карликовых планетах и кометах в Солнечной системе, без упоминания биологии.
Наконец, следует обратиться к идее “некропанспермии” (Wesson, 2011) - идее о том, что организмы постоянно прибывают на Землю, но настолько разлагаются под воздействием ионизирующего излучения и обезвоживаются из-за суровых условий космоса, что поступают на Землю в мертвом состоянии. Тем не менее, они пополняют земной генофонд (ДНК/РНК), являясь даже мертвыми. Мы рассмотрели это в более раннем варианте, но решили не включать его в основной раздел. Тогда мы думали (Wickramasinghe, 2011) и до сих пор думаем, что идеи Вессона противоречивы и, следовательно, нежизнеспособны с научной точки зрения в широком диапазоне всех доказательств продолжающегося прибытия живых организмов на Землю (со свойственными им выносливостью и резистентностью к космическим условиям на протяжении последних 4 миллиардов лет).
Приложение В. Ретровирусы, ретроэлементы Alu и LINE: структура, функции, подвижность генома и значение
Диспергированные короткие (SINE) и длинные (LINE) повторы ретроэлементов (RLE) в геноме человека и млекопитающих являются основными характеристиками структуры генома и движущими силами геномного эволюционного разнообразия (см. Обзоры Kapitonov and Jurka, 1996; Arcot et al., 1996; Cordaux and Batzer, 2009) и ретротранспозиции зародышевой линии этого типа, которые обсуждались намного раньше
40 Заново (Прим.пер.)
41 На месте (Прим.пер.)
как общие эволюционные механизмы в работе Стила (Steele et al., 1984). Они в большом количестве распределены по геному посредством ретротранспозиции через обратную транскриптазу. LINE RLE - это ядерные элементы c длинными вкраплениями (около 500000 пар оснований в геноме человека), при этом различные группы элементов были постоянно активными (в их геномной мобильности) на протяжении последних 150 млн. лет, а текущая активность основных элементов L1 имеет важное значение для функционирования мозга и ЦНС здорового человека, выполняя особые нейронные функции. (Erwin et al., 2016). Элементы L1 имеют размер около 6 КБ, кодируют свою собственную обратную транскриптазу (RT)42 и обеспечивают эту активность ревертазы для совместной мобилизации более мелких элементов Alu (длиной около 300 п.н., общее число которых составляет примерно 1,1 миллиона копий в геноме человека). LINE и Alu RLE расположены в транскрипционно активных насыщенных генами областях генома, обычно в интронах (поскольку их вставка в кодирующие области, когда это случается, обычно приводит к таким драматическим нарушениям генов, что мощная система выведения очищает организм от таких летальных мутаций, допуская только безвредные и “нейтральные” вставки LINE и Alu, которые обычно лежат вне участков, кодирующих белок). Однако ретроэлементы LINE и Alu имеют функционально-важное значение (Steele, 2015; Erwin et al., 2016) с повторами Alu, локализованными в интронных областях нейронов мозга и ЦНС, которые имеют большую значимость (Paz-Yaacov et al., 2010). Элементы Alu имеют димерную структуру, первоначально полученную из РНК 7SL РНК частицы, распознающей сигнал гена, и в их центральных частях они насыщены аденозином (A) и, таким образом, являются идеальными мишенями в структурах дцРНК для пре-мРНК редактирования, превращающим аденозин в инозин (A-to-I), опосредованного дезаминазами ADAR, - опять же выраженная активность, имеющая важное значение для мозга и ЦНС человека и высших приматов (Paz-Yaacov et al., 2010). Действительно, “Alu-экзонизация” или создание нового гена посредством редактирования РНК с переходом к новым сайтам сплайсинга пре-мРНК (таким образом, пропуск экзонов во время сплайсинга пре-мРНК) одновременно генерирует и разрушает существующие экзоны и, таким образом, создает новые изоформы генов (на которых физиологический соматический отбор может действовать для усиления синаптических функций мозга и ЦНС). Такая перестройка генов соматических нейронов может, конечно, также привести к патологии и заболеваниям, таким как болезнь Альцгеймера, болезнь Паркинсона, расстройство аутистического спектра. Установленные семейства Alu повторов являются очень хорошими эволюционными маркерами. Учитывая установленные последовательности, их можно разделить как минимум на три подсемейства (и их под-подсемейства). Самый старый Alu J, расширенный за счет транскрипции или обратной транскрипции 81 млн лет назад; характерные для приматов Alu S, под-подсемейства Sx, Sq, Sp, Sc, - все характерные для платирринов, распространены по геному 35-44 млн.лет назад; и Sg - около 31 млн лет назад. Характерное для приматов подсемейство Alu Y распространилось от 19 млн лет назад до всего нескольких сотен тысяч лет назад. Такие маркеры, связанные с другими появляющимися новыми генами, позволяют производить молекулярное “датирование” появления новых семейств генов зародышевой линии в линиях приматов, ведущих к Homo Sapiens. Наряду с аналогичными эндогенными ретровирусами (ERV, около 10 Кб и > 200 000 пар на гаплоидный геном человека), множественные пары оснований ретроэлементов LINE и Alu являются движущими силами
42 От англ. Reverse Transcriptase (Прим.пер.)
дублирующих геномных расширений, поскольку их аналогичные последовательности, разделенные, но функционирующие в физическом тандеме, могут управлять большими сегментарными дупликациями за счет несовпадающей рекомбинации во время мейоза, тем самым увеличивая как размер генома, так и функциональное разнообразие (Dawkins et al., 1999; McLure et al., 2013; Steele, 2014, 2015).
Особенностью ретровирусов является их способность включать случайно выбранные ферменты клетки-хозяина и малые РНК различных типов в вирионы. А также белки обратной транскриптазы (RT) (> 50 на вирион) и полученные от хозяина ферменты APOBEC3G, которые являются мутаторами ДНК (дезаминирование C-to-U оцДНК), и специфичные для хозяина пре-тРНК, которые инициируют обратную транскрипцию. Отдельная упаковка множественных некодирующих РНК клетки-хозяина также включает РНК 7SL-РНК частицы распознавания сигнала (300-нуклеотидные предшественники РНК ретроэлементов Alu-РНК), а также сами РНК Alu-хозяина, специфические регуляторные микроРНК и сплайсосомную малую ядерную РНК (мяРНК) U6, которая присутствует, как предполагается, только в ядрах млекопитающих, что указывает на новые механизмы вовлечения этих РНК в сборку ретровирусов и их вирулентность. Таким образом, хотя два вирусных генома размером 10 т.п.н.43 являются основными обитателями зрелых ретровирусных вирионов, в настоящее время признано, что заражающий вирус доставляет гораздо больше дополнительных специфических белков и видов нуклеиновых кислот к намеченной клетке-мишени, чем считалось ранее (Tian et al., 2007; Bach, 2008; Hu et al., 2012; Lin et al., 2012; Eckwahl et al., 2016; Telesnitsky et al., 2016), - что, без сомнения, помогает очень мутагенной клеточной среде, вызванной инфицированием ВИЧ и другими вирусами.
В тексте упоминается “Гипотеза транспонируемого элемента (TЭ) тяги” Оливера и Грина (Oliver and Greene, 2012), которая рассматривает внутреннюю силу, периодически подталкивающую к адаптированию, видообразованию и новаторской эволюции часто, но не всегда, выверенным равновесным образом. Когда большинство TЭ деактивируются накопленными мутациями, и если они не заменяются горизонтальным переносом транспозонов или эндогенизированными ретровирусами и т. д., клоны могут стать “живыми ископаемыми", и, кроме того, если этот недостаток транспонируемого элемента тяги сохранится, то линии могут исчезнуть как часть фонового вымирания, которое, по-видимому, составляет подавляющее большинство вымираний на Земле.
Приложение С. Анализ причин и движущих сил истории происхождения жизни на Земле, изложенных в этой статье
По словам соавтора этой статьи Джона А. Шустера, доктора исторических и философских наук, специалиста по Декарту и Коперниканской революции, защитившего докторскую диссертацию под руководством Томаса С. Куна: “Вышеупомянутая статья представляет собой радикальный вызов нынешнему научному консенсусу относительно причин, которые используются для объяснения истории жизни на Земле. Принятые в настоящее
43 Тысяча пар нуклеотидов (прим.пер.)
время дарвиновские механизмы, связанные с передовым опытом по исследованию геологической истории Земли, не подвергаются сомнению. Однако они помещены в новый набор взаимозависимостей с более широкой теоретической цепочкой событий, основанной, с одной стороны, на развитии астрофизики, а затем астробиологии, начиная с первоначальных вкладов и текущей работы Фреда Хойла и Н. Чандры Викрамасингхе и их коллег и, с другой стороны, - на разработках вирусологии и молекулярной биологии, включая, что наиболее важно, постоянные усовершенствования неоламаркианских моделей в молекулярной иммунологии, разработанные Эдвардом Дж. Стилом и рядом коллег за последние сорок лет.
Первоначально можно представить себе природу и объем нового синтеза по аналогии с тем, как дифференциальные уравнения, описывающие известные законы, взаимодействуют с граничными условиями, которые необходимо ввести в них для получения конкретных решений. Можно сказать, что обычно согласованные дарвиновские процессы, наряду с неоламаркистскими процессами Стила, всегда действовали на Земле. Они аналогичны обычным уравнениям или алгоритмам, используемым для описания системы. Что касается граничных условий, применяемых к уравнениям для получения конкретных решений, у нас теперь есть радикально новое видение. Есть перемещающиеся, непредсказуемые и, с геологической точки зрения, внезапные астробиологические воздействия. Как будто кто-то время от времени настаивал на случайном изменении граничных условий. История жизни на Земле становится временной записью результатов основных дарвиновских/неоламаркистских механизмов или уравнений, поскольку они работали сверхурочно над конкатенацией случайно заданных по времени, быстро и радикально измененных граничных условий. Во времена новых знаний о космической биологии состояние жизни на Земле станет результатом всей предыдущей работы фундаментальных механизмов на последовательной основе предыдущих астробиологических данных. С поступлением новой информации система продолжит свое развитие, основываясь на знаниях, механизмах и предыдущих достижениях.
Как утверждается в статье, причинные элементы, вовлеченные в биологическую эволюцию, должны радикально измениться за пределами неодарвинистского консенсуса. Это достигается за счет понимания и концептуального изложения друг другу результатов и моделей из ранее весьма разрозненных областей, таких как астробиология и молекулярная иммунология. Новый синтез предполагает, что конкретная история жизни на Земле обязательно откроет факты о биологических группировках и сопоставления между ними, значительно отличающиеся от тех, которые были наложены при чтении доказательств исключительно в свете неодарвинизма. Подобно тому, как первоначальное видение Николая Коперника в 1543 году было полностью воспринято лишь несколькими поколениями позже на основе введения Кеплером законов и возможных причинных механизмов движения планет и открытия Галилеем убедительных подтверждающих эмпирических данных, так и модель панспермии Хойла-Викрамасингхе теперь можно конкретизировать и сформулировать с помощью двух поколений открытий в астробиологии и молекулярной биологии. В каждом случае первоначальные утверждения были потенциально революционными и сами по себе были блестяще реализованы. Но в каждом случае необходимо было заложить более широкий и глубокий эмпирический и теоретический фундамент под первоначально заявленные основы с использованием
фактических и теоретических материалов, которые не всегда были доступны во время инаугурационных требований основателей.
Наконец, давайте взглянем еще шире и поразмышляем о проблемах, которые новый синтез ставит перед целым рядом политических, культурных и религиозных убеждений. Революционные теории почти всегда имеют такую полутень более широких последствий. В документе на это намекают, конечно, без возможности вдаваться в какие-либо подробности. Например, среди более широкой научно грамотной общественности под угрозой находятся доминирующие популярные версии неодарвинизма, а также многие культурные, философские и общественно-политические утверждения, которые были извлечены из него. Для верующих всех мастей новый синтез, даже в большей степени, чем ортодоксальный дарвинизм, демистифицирует историю жизни на Земле и включает ее в историю космических событий, то есть в историю Вселенной в целом. Наконец, как давно признали историки и историки науки, существует главный вопрос, который с начала 19 века подпитывал как биологические, так и социально-политические теории, то есть даже до Дарвина. Этот вопрос касается “места человека в природе”, где “природа” означает “непосредственное земное окружение”. Новый синтез заново изобретает и расширяет этот вопрос: теперь мы будем спрашивать о месте человечества и отношениях со всем космосом, и, в частности, мы будем рассматривать положение человеческого рода как результат конкатенации космических процессов и событий, которые повлияли на Землю явно разорванным и случайно-причинным образом.” (Примечание: физиолог-эволюционист Денис Нобл из Оксфорда также использовал доказательство, используя понятия граничных и начальных условий (глава 6 “Танец под мелодию жизни”, 2017, и до этого были использованы им же в работе 2011 года).
- Aaskov, J., et al., 2006. Long- term transmission of defective RNA viruses in humans and Aedes mosquitoes. Science 311, 236-238.
- Abe, F., et al., 2013. Extending the planetary mass function to Earth mass by microlensing at moderately high magnification. Mon. Not. Roy. Astron. Soc. 431 (4), 2975-2985.
- Aiewsakun, P., Katzourakis, A., 2017. Marine origin of retroviruses in the early Palaeozoic Era. Nat. Commun. 8, 13954. https://doi.org/10.1038/ncomms13954.
- Albertin, C.B., et al., 2015. The octopus genome and the evolution of cephalopod neural and morphological novelties. Nature 524, 220-224.
- Allen, D.A., Wickramasinghe, D.T., 1981. Diffuse interstellar absorption bands between 2.9 and 4.0 mm. Nature 294, 239-240.
- Altwegg, K., et al., 2016. Prebiotic chemical - amino acid and phosphorus in the coma of comet 67P/Churyumov-Gerasimenko. Sci. Adv 2, e1600285, 27 May.
- Arcot, S.S., et al., 1996. Alu fossil relics - distribution and insertion polymorphism. Genome Res. 6, 1084-1092.
- Bach, D., 2008. Characterization of APOBEC3G binding to 7SL RNA. Retrovirology 5, 54. https://doi.org/10.1186/1742-4690-5-54.
- Bell, E.A., et al., 2015. Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon. Proc. Natl. Acad. Sci. U.S.A. 112, 14518-14521.
- Bergmann, S., et al., 2006. The hemocyanin from a living fossil, the cephalopod Nautilus pompilius: protein structure, gene organization, and evolution. J. Mol. Evol. 62, 362-374.
- Bianciardi, G., et al., 2012. Complexity analysis of the viking labelled Release experiments. Intl J. Aeronautical & Space Sci. 13, 14-26. https://doi.org/10.5139/ IJASS.2012.13.1.14.
- Bieler, K., et al., 2015. Abundant molecular oxygen in the coma of comet 67P/ChuryumoveGerasimenko. Nature 526, 678-681. https://doi.org/10.1038/ nature15707.
- Biver, N., et al., 2015. Ethyl alcohol and sugar in comet C/2014 Q2 (Lovejoy). Sci. Adv. 1, e1500863, 23 October 2015.
- Bostrom, N., 2014. Superintelligence: Paths, Dangers, Strategies. Oxford University Press, Oxford, UK.
- Brasier, M., 2012. Secret Chambers. Oxford University Press. ISBN 9780199644001.
- Buhr, F., et al., 2016. Synonymous codons direct co-translational folding towards different protein conformations. Mol. Cell 61, 342-351.
- Campbell, J.H., Perkins, P., 1988. Transgenerational effects of drug and hormonal treatments in mammals: a review of observations and ideas. In: Boer, G.J., Feenstra, M.G.P., Mirmiran, M., Swaab, D.F., van Haaren, F. (Eds.), Progress in Brain Research, vol. 73, pp. 535-553.
- Capaccione, F., et al., 2015. The organic-rich surface of comet 67P/ChuryumovGerasimenko as seen by VIRTIS/Rosetta. Science 347 (6220).
- Carlini, D.B., et al., 2000. Actin gene family evolution and the phylogeny of coleoid cephalopods (Mollusca: cephalopoda). Mol. Biol. Evol. 17, 1353-1370.
- Chen, Y.-H., et al., 2015. Phosphatidylserine vesicles enable efficient en bloc transmission of enteroviruses. Cell 160, 619-630. https://doi.org/10.1016/ j.cell.2015.01.032.
- Chiu, Y.-L., Greene, W.C., 2006. APOBEC3 Cytidine Deaminases: distinct antiviral actions along the retroviral life cycle. J. Biol. Chem. 281, 8309-8312. https:// doi.org/10.1074/jbc.R500021200.
- Chivian, D., et al., 2008. Environmental genomics reveals a single-species ecosystem deep within the Earth. Science 322, 275-278.
- Christener, B.C., et al., 2014. A microbial ecosystem beneath the West Antarctic ice sheet. Nature 512, 310-313. https://doi.org/10.1038/nature13667.
- Clube, S.V.M., et al., 1996. Giant comets, evolution and civilization. Astrophys. Space Sci. 245, 43-60.
- Combe, M., et al., 2015. Single-cell analysis of RNA virus infection identifies multiple genetically diverse viral genomes within single infectious units. Cell Host Microbe 18, 424-432. https://doi.org/10.1016/j.chom.2015.09.009.
- Cordaux, R., Batzer, M.A., 2009. The impact of retrotranspositions on human genome evolution. Nat. Rev. Genet. 10, 691-703.
- Cosic, I., 1994. Macromolecular bioactivity: is it resonant interaction between macromolecules? - Theory and applications. IEEE Trans. Biomed. Eng. 41, 1101-1114.
- Cosic, I., Cosic, D., 2016. The treatment of crigler-najjar syndrome by blue light as explained by resonant recognition model. EPJ Nonlinear Biomedical Physics 4, 9. https://doi.org/10.1140/epjnbp/s40366-016-0036-6.
- Cosic, I., et al., 2015. Is it possible to predict electromagnetic resonances in proteins, DNA and RNA? EPJ Nonlinear Biomedical Physics 3, 5. https://doi.org/10.1140/ s40366-015-0020-6.
- Cosic, I., et al., 2016. Environmental light and its relationship with electromagnetic resonances of biomolecular interactions, as predicted by the Resonant Recognition Model. Int. J. Environ. Res. Publ. Health 13 (7), 647,. https://doi.org/ 10.3390/ijeprh13070647.
- Cossetti, C., et al., 2014. Soma-to- germline transmission of RNA in mice xenografted with human tumour cells: possible transport by exosomes. PLoS One 9, e101629.
- Coulson, S.G., Wickramasinghe, N.C., 2000. IR spectrometry of coals. In: Celnikier, L.M., Tran Thanh Van, J. (Eds.), Frontiers of Life, Proceedings of the XIIth Recontres de Blois, pp. 233-236.
- Coulson, S.G., Wickramasinghe, N.C., 2003. Frictional and radiation heating of micron-sized meteoroids in the Earth's upper atmosphere. Mon. Not. Roy. Astron. Soc. 343, 1123-1130.
- Coulson, S.G., et al., 2014. On the dynamics of volatile meteorites. Mon. Not. Roy. Astron. Soc. 445, 3669-3673.
- Crick, F.H.C., Orgel, L.E., 1973. Directed panspermia. Icarus 19, 341-346.
- Dawkins, R.L., 2015. Adapting Genetics: Quantal Evolution after Natural Selection e Surviving the Changes to Come. Nearurban Publishing, Dallas, TX. ISBN 978-0- 9864115-1-9.
- Dawkins, R.L., et al., 1999. Genomics of the major histocompatibility complex: haplotypes, duplication, retroviruses and disease. Immunol. Rev. 167, 275-304.
- Deamer, D., 2011. First Life: Discovering the Connections between Stars, Cells, and How Life Began. University of California Press, Berkeley, USA.
- Devanapally, S., et al., 2015. Double-stranded RNA made in C. elegans neurons can enter the germline and cause transgenerational gene silencing. Proc. Natl. Acad. Sci. U.S.A. 112, 2133-2138. https://doi.org/10.1073/pnas.1423333112.
- Diehl, W.E., et al., 2016. Tracking interspecies transmission and long-term evolution of an ancient retrovirus using the genomes of modern mammals. eLife 5, e12704.
- Dodd, M.C., et al., 2017. Evidence for early life in Earth's oldest hydrothermal vent precipitates. Nature 543, 60-65.
- Doria, M., et al., 2009. Editing of HIV-1 RNA by the double-stranded RNA deaminase ADAR1 stimulates viral infection. Nucleic Acids Res. 37, 5848-5858.
- Eckwahl, M.J., et al., 2016. Host RNA packaging by retroviruses: a newly synthesized story. mBio 7 (1). https://doi.org/10.1128/mBio.02025-15 e02025-15.
- Eldredge, N., Gould, S.J., 1972. Punctuated equilibria : an alternative to phyletic gradualism. In: Schopf, T.J.M. (Ed.), Models in Paleobiology. Freeman Cooper, San Francisco, pp. 82-115.
- Erwin, J.A., et al., 2016. L1-associated genomic regions are deleted in somatic cells of the healthy human brain. Nat. Neurosci. 19, 1583-1591. https://doi.org/10.1038/ nn.4388.
- Flew, A., 2007. There Is a God : How the World's Most Notorious Atheist Chang His is Mind. Harper Collins, New York.
- Fox, D., 2014. Antartica's secret garden. Nature 512, 244-246.
- Frank, L.A., Sigwarth, J.B., 2001. Detection of small comets with a ground-based telescope. J. Geophys. Res. 106, 3665-3683.
- Franklin, A., et al., 2004. Human DNA polymerase-h, an A-T mutator in somatic hypermutation of rearranged immunoglobulin genes, is a reverse transcriptase. Immunol. Cell Biol. 82, 219-225. https://doi.org/10.1046/j.0818- 9641.2004.01221.x.
- Gibson, C., et al., 2011. The origin of life from primordial planets. Int. J. Astrobiol. 10, 83e98. https://doi.org/10.1017/S1473550410000352. https://arxiv.org/abs/ 1004.0504.
- Gillon, et al., 2016. Temperate earth size planets transiting a nearby ultracool dwarf star. Nature 533, 221-224.
- Gold, T., 1989. New ideas in science. J. Sci. Explor. 3 (2), 103-112. http://amasci.com/ freenrg/newidea1.html.
- Gold, T., 1992. The deep hot biosphere. Proc. Natl. Acad. Sci. U.S.A. 89, 6045-6049.
- Gold, T., 1999. The Deep Hot Biosphere: the Myth of Fossil Fuels. Copernicus. Springer Verlag, New York.
- Gorczynski, R.M., Steele, E.J., 1980. Inheritance of acquired immunologic tolerance to foreign histocompatibility antigens in mice. Proc. Natl. Acad. Sci. U.S.A. 77, 2871-2875.
- Gorczynski, R.M., Steele, E.J., 1981. Simultaneous yet independent inheritance of somatically acquired tolerance to two distinct H-2 antigenic haplotype determinants in mice. Nature 289, 678-681. https://doi.org/10.1038/289678a0.
- Gould, S.J., Eldredge, N., 1977. Punctuated equilibria: the tempo and mode of evolution reconsidered. Paleobiology 3, 115-151.
- Grebennikova, T.V., Syroeshkin, A.V., Shubralova, E.V., et al., 2018. The DNA of bacteria of the World Ocean and the Earth in cosmic dust at the International Space Station. Sci. World J. In press.
- Harris, M.J., et al., 2002. The detection of living cells in the stratosphere. Proc.SPIE Conf. 4495, 192-198.
- Harris, R.S., Liddament, M.T., 2004. Retroviral restriction by APOBEC proteins. Nat. Rev. Immunol. 4, 868-877.
- Haynes, R., 1980. The boggle threshold. Encounter 92-96. Aug 1980.
- Нiel, C.S., et al., 2014. Functional activity of plasmid DNA after entry into the atmosphere of Earth investigated by a new biomarker stability assay for Ballistic Spaceflight Experiments. e112979 PLoS One 9.
- Hoover, R.B., 2005. Microfossils, biominerals, and chemical biomarkers in meteorites. In: Hoover, R.B., Rozanov, A.Y., Paepe (Eds.), Perspectives in Astrobiology. RR IOS Press, Amsterdam, pp. 43-65.
- Hoover, R.B., 2011. Fossils of cyanobacteria in CI1 carbonaceous meteorites: implications to life on comets, Europa and Enceladus. J. Cosmology 16, 7070-7111.
- Hope-Simpson, R.E., 1978. Sunspots and flu: a correlation. Nature 275, 86.
- Hoyle, F., Wickramasinghe, N.C., 1976. Primitive grain clumps and organic compounds in carbonaceous chondrites. Nature 264, 45.
- Hoyle, F., Wickramasinghe, N.C., 1977a. Polysaccharides and the infrared spectra of galactic sources. Nature 268, 610-612.
- Hoyle, F., Wickramasinghe, N.C., 1977b. Identification of the 2200A interstellar absorption feature. Nature 270, 323.
- Hoyle, F., Wickramasinghe, N.C., 1978a. Calculations of infrared fluxes from galactic sources for a polysaccharide grain model. Astrophys. Space Sci. 53, 489-505.
- Hoyle, F., Wickramasinghe, N.C., 1978b. Life Cloud. J.M. Dent Ltd, London.
- Hoyle, F., Wickramasinghe, N.C., 1979. Diseases from Space. J.M. Dent Ltd, London.
- Hoyle, F., Wickramasinghe, N.C., 1981. Evolution from Space. J.M. Dent Ltd, London.
- Hoyle, F., Wickramasinghe, C., 1982. Why Neo-Darwinism Does Not Work. University College Cardiff Press. ISBN 0 906449 50 2.
- Hoyle, F., Wickramasinghe, N.C., 1985. Living Comets. Univ. College, Cardiff Press, Cardiff.
- Hoyle, F., Wickramasinghe, N.C., 1986. The case for life as a cosmic phenomenon. Nature 322, 509-511.
- Hoyle, F., Wickramasinghe, N.C., 1991. The Theory of Cosmic Grains. Kluwer, Dordrecht.
- Hoyle, F., Wickramasinghe, N.C., 1993. Our Place in the Cosmos : the Unfinished Revolution. J.M. Dent Ltd, London.
- Hoyle, F., Wickramasinghe, N.C., 1997. Life on Mars? : the Case for a Cosmic Heritage. Clinical Press Ltd, Bristol.
- Hoyle, F., Wickramasinghe, N.C., 2000. Astronomical Origins of Life: Steps towards Panspermia. Kluwer, Dordrecht.
- Hoyle, F., et al., 1982. Infrared spectroscopy over the 2.9-3.9mm waveband in biochemistry and astronomy. Astrophys. Space Sci. 83, 405-409.
- Hoyle, F., et al., 1984. The spectroscopic identification of interstellar grains. Astrophys. Space Sci. 98, 343-352.
- Hu, W.-S., et al., 2012. HIV-1 reverse transcription. Cold Spring Harb Perspect Med 2 a006882, 22 pages.
- Jablonka, E., Lamb, M.J., 1995. Epigenetic Inheritance and Evolution:The Lamarckian Dimension. Oxford University Press, Oxford.
- Jablonka, E., Lamb, M.J., 2005. Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History of Life. MIT Press, Boston. ISBN 0-262-10107-6.
- Jones, R.B., et al., 2013. LINE-1 retrotransposable element DNA accumulates in HIV1- infected cells. J. Virol. 87, 13307-13320.
- Jönsson, K.I., et al., 2008. Tardigrades survive exposure to space in low Earth orbit. € Curr. Biol. 18, R729-R731.
- Kapitonov, V., Jurka, J., 1996. The age of Alu subfamilies. J. Mol. Evol. 42, 59-65.
- Katzourakis, A., 2013. Paleovirology: inferring viral evolution from host genome E.J. Steele et al. / Progress in Biophysics and sequence data. Phil. Trans. Roy.Soc. Lond. 368, 20120493
- Kopparapu, R.K., 2013. A revised estimate of the occurrence rate of terrestrial planets in habitable zones around Kepler M-Dwarf. Astrophys. J. 767. L8.
- Kuhn, T.S., 1970. The Structure of Scientific Revolutions, second ed. Univ. Chicago Press, Chicago.
- Lane, N., 2015. The Vital Question: Energy, Evolution, and the Origins of Complex Life. W.W. Norton & Company, London.
- Levin, G.V., 2007. Modern myths of Mars. Int. J. Astrobiol. 6, 95-108.
- Levin, G.V., 2013. Implications of Curiosity's findings for the viking labelled Release experiment and life on Mars. Instruments, methods, and missions for astrobiology XVI. In: SPIE Proc., 8865, 8865-8862.
- Levin, G.V., 2015. The curiousness of Curiosity. Astrobiology 15, 101e103. https:// doi.org/10.1089/ast.2014.1406.
- Levin, G.V., Straat, P.A., 1976. Viking Labeled Release biology experiment: interim results. Science 194, 1322-1329.
- Levin, G.V., Straat, P.A., 2016. The case for extant life on Mars and its possible detection by the Viking Labelled Release Experiment. Astrobiology 16, 798-810.
- Levison, H.F., et al., 2010. Capture of the Sun's Oort cloud from stars in its birth cluster. Science 329, 187-190. https://doi.org/10.1126/science.1187535.
- Leya, T., et al., 2017. Algae Survive outside Space Station. AlgaeIndustryMagazine.Com, 7th February, 2017. http://www.algaeindustrymagazine. com/algae-survive-outside-space-station/.
- Liscovitch-Brauer, et al., 2017. Trade-off between transcriptome plasticity and genome evolution in cephalopods. Cell 169, 191-202.
- Lin, X., 2012. MicroRNAs and unusual small RNAs discovered in Kaposi's SarcomaAssociated Herpesvirus virions. J. Virol. 86, 12717-12730. https://doi.org/ 10.1128/JVI.01473-12.
- Lindley, R., 2010. The Soma: How Our Genes Really Work and How that Changes Everything! CYO Foundation. ISBN1451525648, POD book Amazon.com.
- Lindley, R., 2013. The importance of codon context for understanding the Ig-like somatic hypermutation strand-biased patterns in TP53 mutations in breast cancer. Cancer Genet. 206, 222-226.
- Lindley, R.A., Steele, E.J., 2013. Critical analysis of strand-biased somatic mutation signatures in TP53 versus Ig genes. In: Genome -wide Data and the Etiology of Cancer ISRN Genomics, vol. 2013. Article ID 921418, 18 pages.
- Lindley, R.A., et al., 2016. Association between targeted somatic mutation (TSM) signatures and HGS-OvCa progression. Cancer Med 5, 2629-2640.
- Liu, Y., 2007. Like father like son. EMBO Rep. 8, 798-803.
- Liu, Y., Li, X., 2016a. Darwin's Pangenesis as a molecular theory of inherited diseases. Gene 582, 19-22. https://doi.org/10.1016/j.gene.2016.01.051.
- Liu, Y., Li, X., 2016b. Darwin and Mendel today: a comment on "Limits of imagination: the 150th Anniversary of Mendel's Laws, and why Mendel failed to see the importance of his discovery for Darwin's theory of evolution". Genome 59, 75-77. https://doi.org/10.1139/gen-2015-0155.
- Louis, G., Kumar, A.S., 2006. The red rain phenomenon in Kerala and its possible extraterrestrial origin. Astrophys. Sp.Sci. 302, 175-187.
- Lutz, F.E., Grisewood, E.N., 1934. Reactions of Drosophila to 2537A radiation. Am. Mus. Novit. 706. March 1934.
- Mattick, J.S., 2012. Rocking the foundations of molecular genetics. Proc. Natl. Acad. Sci. U. S. A. 109, 16400-16401.
- McCafferty, P., 2008. Bloody red rain again! Red rain in meteors and myth. Int. J. Astrobiol. 7, 9-15.
- McLure, C.A., et al., 2013. Genomic evolution and polymorphism: segmental duplications and haplotypes at 108 regions on 21 chromosomes. Genomics 102, 15-26.
- Miyake, N., et al., 2010. Identification of micro-biofossils in space dust. J. Cosmology 7, 1743-1749.
- Muotri, A.R., et al., 2007. The necessary junk: new functions for transposable elements. Hum. Mol. Genet. 16 (2), R159-R167. https://doi.org/10.1093/hmg/ ddm196.
- Napier, W.M., et al., 2007. The origin of life in comets. Int. J. Astrobiol. 6, 321-323.
- Noble, D., 2011. A Theory of Biological Relativity: No Privileged Level of Causation Interface Focus Royal Society Publishing. https://doi.org/10.1098/rsfs.2011.0067. Published 9 November 2011.
- Noble, D., 2013. Physiology is rocking the foundations of evolutionary biology. Exp. Physiol. 98 (8), 1235-1243.
- Noble, D., 2017. Dance to the Tune of Life. Cambridge University Press. ISBN 1-107- 17624-9.
- Noble, D., et al., 2016. New Trends in Evolutionary Biology: Biological, Philosophical and Social Science Perspectives. The Royal Society, London, 6-9 Carlton House Terrace, London, SW1Y 5AG Nov 7 -9, 2016. Editors-Organisors. https:// royalsociety.org/science-events-and-lectures/2016/11/evolutionary-biology/.
- Oliver, K.R., Greene, W.K., 2012. Transposable elements and viruses as factors in adaptation and evolution: an expansion and strengthening of the TE-Thrust hypothesis. Ecol. Evol. 2 (11), 2912-2933. https://doi.org/10.1002/ece3.400.
- Paules, C.I., et al., 2017. The pathway to a universal influenza vaccine. Immunity 47, 599-603.
- Paz-Yaacov, N., et al., 2010. Adenosine-to-inosine RNA editing shapes transcriptome diversity in primates. Proc. Natl. Acad. Sci. U.S.A. 107, 12174-12179.
- Pflug, H.D., Heinz, B., 1997. Analysis of fossil organic nanostructures: terrestrial and extraterrestrial. SPIE Proc. Instrum. Methods Missions Investigation Extraterr. Microorg.. 86, 3111. https://doi.org/10.1117/12.278814. July 11, 1997.
- Pirakitikulr, N., et al., 2016. The coding region of the HCV genome contains a network of regulatory RNA structures. Mol. Cell 61, 1-10
- Priscu, J.C., Christner, B.C., 2004. Earth's icy biosphere. In: Bull, A. (Ed.), Microbial Diversity and Bioprospecting. ASM Press, Washington, D.C., pp. 130-145. Chap 13.
- Qu, J., 2016. Is sunspot activity a factor in influenza pandemics? Rev. Med. Virol. 26, 309-313. https://doi.org/10.1002/rmv.1887.
- Rauf, K., Wickramasinghe, C., 2010. Evidence for biodegradation products in the interstellar medium. Int. J. Astrobiol. 9, 29-34.
- Refsland, E.W., Harris, R.S., 2013. The APOBEC3 family of retroelement restriction factors. Curr. Top. Microbiol. Immunol. 371, 1-27. https://doi.org/10.1007/978- 3-642-37765-5_1.
- Rothenfluh, H.S., 1995. Hypothesis: a memory lymphocyte specific soma-togermline genetic feedback loop. Immunol. Cell Biol. 73, 174-180.
- Ruff, S.W., Farmer, J.D., 2016. Silica deposits on Mars with features resembling hot spring biosignatures at El Tatio in Chile Nat. Commun. Now. 7, 13554. https://doi.org/10.1038/ncomms13554.
- Ryan, F., 2009. Virolution. Collins, London.
- Sanjuan, R., 2010. Mutational fitness effects in RNA and single-stranded DNA viruses: common patterns revealed by site-directed mutagenesis studies. Phil Trans R Soc B 365, 1975-1982. https://doi.org/10.1098/rstb.2010.0063.
- Sanjuan, R., 2012. From molecular genetics to phylodynamics: evolutionary relevance of mutation rates across viruses. PLoS Pathog. 8 (5), e1002685. https:// doi.org/10.1371/journal.ppat.1002685.
- Sanjuan, R., Domingo-Calap, P., 2016. Mechanisms of viral mutation. Cell. Mol. Life Sci. 3, 4433e4448. https://doi.org/10.1007/s00018-016-2299-6.
- Sanjuan, R., et al., 2004. The distribution of fitness effects caused by singlenucleotide substitutions in an RNA virus. Proc. Natl. Acad. Sci. U.S.A. 101, 8396-8401. https://doi.org/10.1073/pnas.0400146101.
- Sanjuan, R., et al., 2010. Viral mutation rates. Virology 84, 9733e9748. https:// doi.org/10.1128/JVI.00694-10.
- Sharma, U., et al., 2015. Biogenesis and function of tRNA fragments during sperm maturation and fertilization in mammals. Science 351, 391-396.
- Shi, M., et al., 2016. Redefining the invertebrate RNA virosphere. Nature 540, 539-543. https://doi.org/10.1038/nature20167.
- Shivaji, S., Chaturvedi, P., Begum, Z., et al., 2009. Janibacter hoylei sp. nov., Bacillus isronensis sp. nov. and Bacillus aryabhattai sp. nov., isolated from cryotubes used for collecting air from the upper atmosphere. Int. J. Syst. Evol. Microbiol. 59, 2977-2986.
- Smith, W.E., September 26, 2013. 2013. Life is a cosmic phenomenon: the “Search for Water” evolves into the “Search for Life”. In: Hoover, R.B., Levin, G.V., Rozanov, A.Y., Wickramasinghe, N.C. (Eds.), Proc. SPIE 8865, Instruments, Methods, and Missions for Astrobiology XVI. San Diego, California, United States. https://doi.org/10.1117/12.2046862.
- Snyder, D.P., 2015. The origins of megacryometeors: troposphere or extraterrestrial? Cosmology 19, 70-86 (Cosmology.com).
- Spadafora, C., 2008. Sperm-mediated “reverse” gene transfer: a role of reverse transcriptase in the generation of new genetic information. Hum. Reprod. 23, 735-740. https://doi.org/10.1093/humrep/dem425.
- Steele, E.J., 1979. Somatic Selection and Adaptive Evolution : on the Inheritance of Acquired Characters, first ed. University of Chicago Press, Chicago. WilliamsWallace, Toronto, 1979; 2nd Edit.
- Steele, E.J., 2014. Reflections on ancestral haplotypes: medical genomics, evolution and human individuality. Perspect. Biol. Med. 57, 179-197.
- Steele, E.J., 2015. Ancestral Haplotypes: Our Genomes Have Been Shaped in the Deep Past. Nearurban Dallas,Tx, ISBN 978-0-9864115-0-2.
- Steele, E.J., 2016a. Origin of congenital defects: stable inheritance through the male line via maternal antibodies specific for eye lens antigens inducing autoimmune eye defects in developing rabbits in utero. In: Levin, M., Adams, D.S. (Eds.), Ahead of the Curve -Hidden Breakthroughs in the Biosciences Chapter 3. Michael Levin and Dany Spencer Adams IOP Publishing Ltd 2016, Bristol, UK.
- Steele, E.J., 2016b. Somatic hypermutation in immunity and cancer: critical analysis of strand-biased and codon-context mutation signatures. DNA Repair 45, 1-24. https://doi.org/10.1016/j.dnarep.2016.07.001.
- Steele, E.J., Dawkins, R.L., 2016. New Theory of HIV Diversification : Why it May Never Be Possible to Make a Protective Vaccine viXra.org 1612.0346. http:// viXra.org/abs/1612.0346.
- Steele, E.J., Lindley, R.A., 2017. ADAR deaminase A-to-I editing of DNA and RNA moieties of RNA: DNA hybrids has implications for the mechanism of Ig somatic hypermutation. DNA Repair 55, 1-6.
- Steele, E.J., Lloyd, S.S., 2015. Soma-to-germline feedback is implied by the extreme polymorphism at IGHV relative to MHC. Bioessays 37, 557-569.
- Steele, E.J., Gorczynski, R.M., Pollard, J.W., 1984. The somatic selection of acquired characters. In: Pollard, J.W. (Ed.), Evolutionary Theory: Paths into the Future. John Wiley, London, pp. 217-237.
- Steele, E.J., Lindley, R.A., Blanden, R.V., 1998. In: Davies, Paul (Ed.), Lamarck's Signature : How Retrogenes Are Changing Darwin's Natural Selection Paradigm. Allen & Unwin, Frontiers of Science: Series. Sydney, Australia, 1998.
- Strugnell, J., Nishiguchi, M.K., 2007. Molecular phylogeny of coleoid cephalopods (Mollusca: cephalopoda) inferred from three mitochondrial and six nuclear loci: a comparison of alignment, implied alignment and analysis methods. J. Molluscan Stud. 73, 399-410.
- Strugnell, J., et al., 2005. Molecular phylogeny of coleoid cephalopods (Mollusca: cephalopoda) using a multigene approach; the effect of data partitioning on resolving phylogenies in a Bayesian framework. Mol. Phylogenet. Evol. 37, 426-441.
- Strugnell, J., et al., 2006. Divergence time estimates for major cephalopod groups: evidence from multiple genes. Cladistics 22, 89-96.
- Telesnitsky, A., Wolin, S.L., 2016. The host RNAs in retroviral articles. Viruses 8,
235. https://doi.org/10.3390/v8080235, 15 pages. Temin, H.M., 1974. On the origin of the RNA tumor viruses. Annu. Rev. Genet. 8, 155-177.
- Tepfer, D., Leach, S., 2017. Survival and DNA damage in plant seeds exposed for 558 and 682 Days outside the international space station. Astrobiology 17, 205-215.
- Tegmark, M., 2014. Our Mathematical Universe. Random House, New York.
- Tian, C., et al., 2007. Virion packaging determinants and reverse transcription of SRP RNA in HIV-1 particles. Nucleic Acids Res. 35, 7288-7302. https://doi.org/ 10.1093/nar/gkm816.
- Vanysek, V., Wickramasinghe, N.C., 1975. Formaldehyde polymers in comets. Astrophys. Space Sci. 33, L19-L28.
- Villarreal, L.P., 2004. Can viruses make us human? Proc. Am. Phil. Soc. 148, 296-323.
- Wainwright, M., Wickramasinghe, N., Narlikar, J., Rajaratnam, P., 2003. Microorganisms cultured from stratospheric air samples obtained at 41km. FEMS Microbiol. Lett. 218, 161-165.
- Wainwright, M., et al., 2014. Recovery of cometary microorganisms from the stratosphere. Astrobiol Outreach 2 (1). https://doi.org/10.4172/2332-2519. 1000110.
- Wainwright, M., et al., 2015a. Biological entities isolated from two stratosphere launches-continued evidence for a space origin J. Astrobiol Outreach 3 (2). https://doi.org/10.4172/2332-2519.1000129.
- Wainwright, M., et al., 2015b. Masses staining positive for DNA-isolated from the stratosphere at a height of 41 km. Astrobiol Outreach 3 (2). https://doi.org/10. 4172/2332-2519.1000130.
- Walker Jr., J., Wickramasinghe, C., 2015. Big Bang and God - an Astro-theology. Palgrave Macmillan, US.
- Wallis, M.K., 2007. Exotic amino acids across the K/T boundary e cometary origin and relevance for species extinction. Int. J. Astrobiol. 6, 303-306.
- Wallis, J., et al., 2013. The Polonnaruwa meteorite - oxygen isotope, crystalline and biological composition. J. Cosmol. 21, 10004-10011.
- Wallis, M.K., Wickramasinghe, N.C., 2015. Rosetta images of Comet 67P/Churyumov-Gerasimenko: inferences from its terrain and structure. J. Astrobiol Outreach 3 (1). https://doi.org/10.4172/2332-2519.1000127.
- Wesson, P., 2011. Necropanspermia. Observatory 131, 63-66.
- Wickramasinghe, N.C., 1974. Formaldehyde polymers in interstellar space. Nature 252, 462-463.
- Wickramasinghe, N.C., 2010. The astrobiological case for our cosmic ancestry. Int. J. Astrobiol. 9, 119-129
- Wickramasinghe, C., 2011. Viva panspermia. Observatory 131, 130-134.
- Wickramasinghe, N.C., 2012. DNA sequencing and predictions of the cosmic theory of life. Astrophys. Space Sci. 343, 1-5.
- Wickramasinghe, N.C., et al., 2012. Life-bearing primordial planets in the solar vicinity. Astrophys. Space Sci 341, 295-299.
- Wickramasinghe, Chandra, 2015a. The Search for Our Cosmic Ancestry. World Scientific, Singapore.
- Wickramasinghe, Chandra, 2015b. Vindication of Cosmic Biology. Tribute to Sir Fred Hoyle (1915-2001). Edit. Chandra Wickramasinghe, World Scientific, Singapore.
- Wickramasinghe, N.C., Rycroft, M., 2018. Transport of submicron dust including bacteria from the Earth’s surface to the high ionosphere. Adv. Astrophys. In press.
- Wickramasinghe, D.T., Allen, D.A., 1986. Discovery of organic grains in Comet Halley. Nature 323, 44-46.
- Wickramasinghe, C., Smith, W.E., 2014. Convergence to panspermia. Hypothesis 12 (1), e9,. https://doi.org/10.5779/hypothesis.v12i1.358.
- Wickramasinghe, N.C., Steele, E.J., 2016. Dangers of adhering to an obsolete paradigm: could Zika virus lead to a reversal of human evolution? J. Astrobiol. Outreach 4 (1). https://doi.org/10.4172/2332-2519.1000147.
- Wickramasinghe, N.C., Tokoro, G., 2014a. Life as a cosmic phenomenon: 1 the socioeconomic control of a scientific paradigm. J. Astrobiol.Outreach 2 (2). https:// doi.org/10.4172/2332-2519.1000113.
- Wickramasinghe, N.C., Tokoro, G., 2014b. Life as a cosmic phenomenon: 2.The panspermia trajectory of Homo sapiens. J. Astrobiol Outreach 2 (2). https://doi. org/10.4172/2332-2519.1000115.
- Wickramasinghe, N.C., et al., 1996. Eruptions of comet hale-bopp at 6.5AU. Astrophys. Space Sci. 240, 161-165.
- Wickramasinghe, J.T., et al., 2009. Liquid water and organics in Comets: implications for exobiology. Int. J. Astrobiol. 8, 281-290.
- Wickramasinghe, J.T., et al., 2010. Comets and the Origin of Life. World Scientific, Singapore.
- Wickramasinghe, N.C., et al., 2015. Rosetta studies of Comet 67P/ChuryumoveGerasimenko: prospects for establishing cometary biology. J. Astrobiol. Outreach 3 (1). https://doi.org/10.4172/2332-2519.1000126.
- Wickramasinghe, N.C., et al., 2017. Sunspot cycle minima and pandemics: the case for vigilance? J. Astrobiol. Outreach 5 (2). https://doi.org/10.4172/2332-2519. 1000159.
- Yohn, C.T., Jiang, Z., McGrath, S.D., Hayden, K.E., Khaitovich, P., et al., 2005. Lineagespecific expansions of retroviral insertions within the genomes of african great apes but not humans and orangutans. PLoS Biol. 3, e110
(Перевод: А. Троян)





{kind=link}